- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- For Contributors
- Policies
- E-Submission

- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- Policies
- For Contributors
Articles
- Page Path
- HOME > J. Microbiol > Volume 63(9); 2025 > Article
-
Full article
Metal ion homeostasis regulates condensin-dependent chromatin architecture and chromosome segregation in Schizosaccharomyces pombe - Seong Ho An, Kyoung-Dong Kim*
-
Journal of Microbiology 2025;63(9):jm.2505008.
DOI: https://doi.org/10.71150/jm.2505008
Published online: August 29, 2025
Department of Systems Biotechnology, Chung-Ang University, Anseong 17546, Republic of Korea
- *Correspondence Kyoung-Dong Kim kdkim0122@cau.ac.kr
© The Author(s), under exclusive licence to Microbiological Society of Korea 2026
This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (CC BY 4.0) (https://creativecommons.org/licenses/by/4.0/) which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
- 2,959 Views
- 92 Download
ABSTRACT
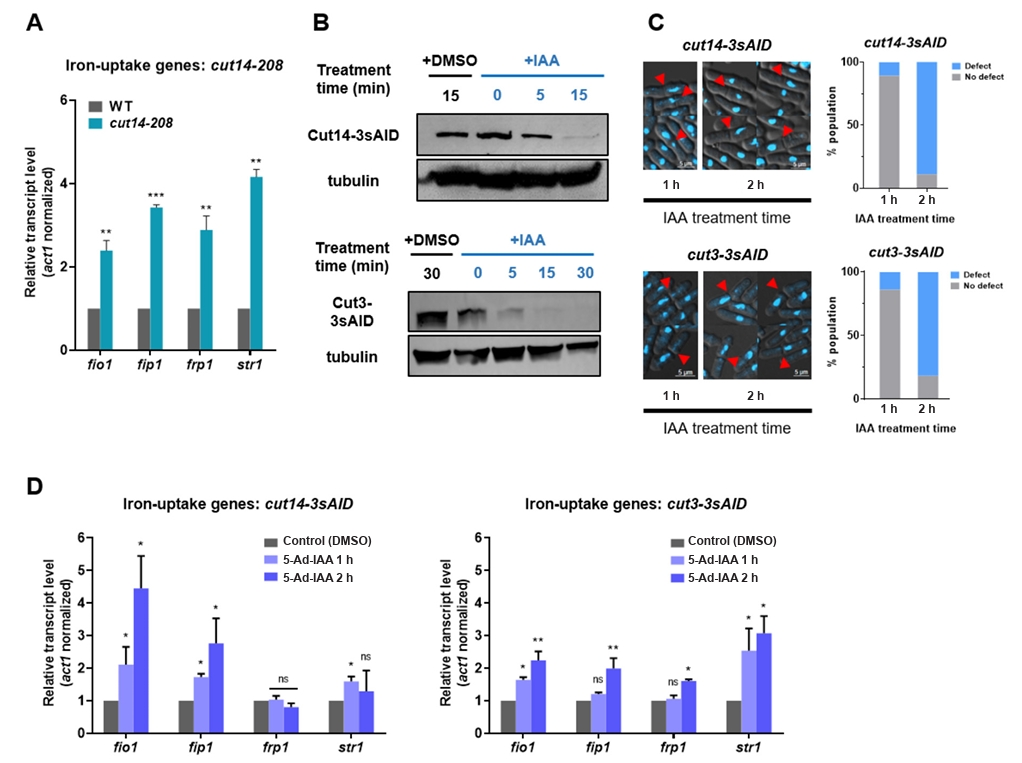
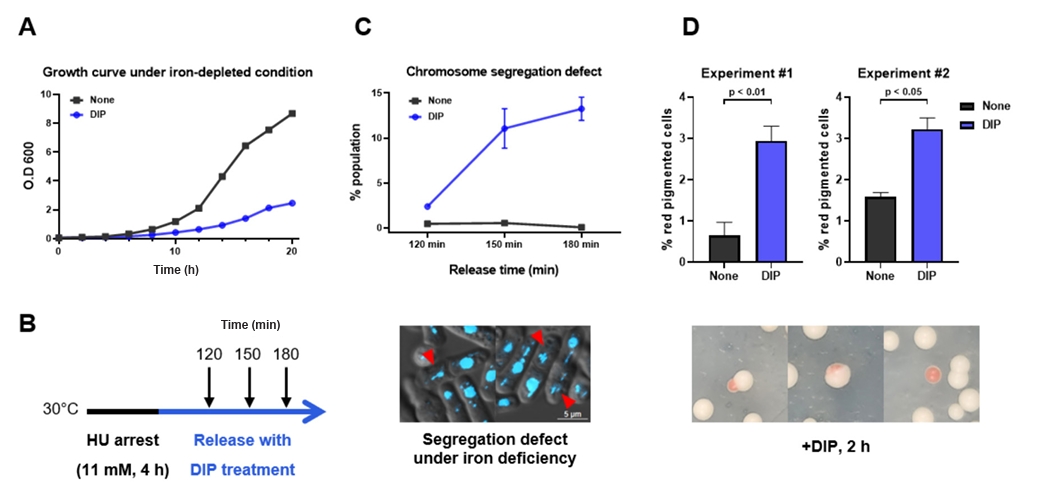
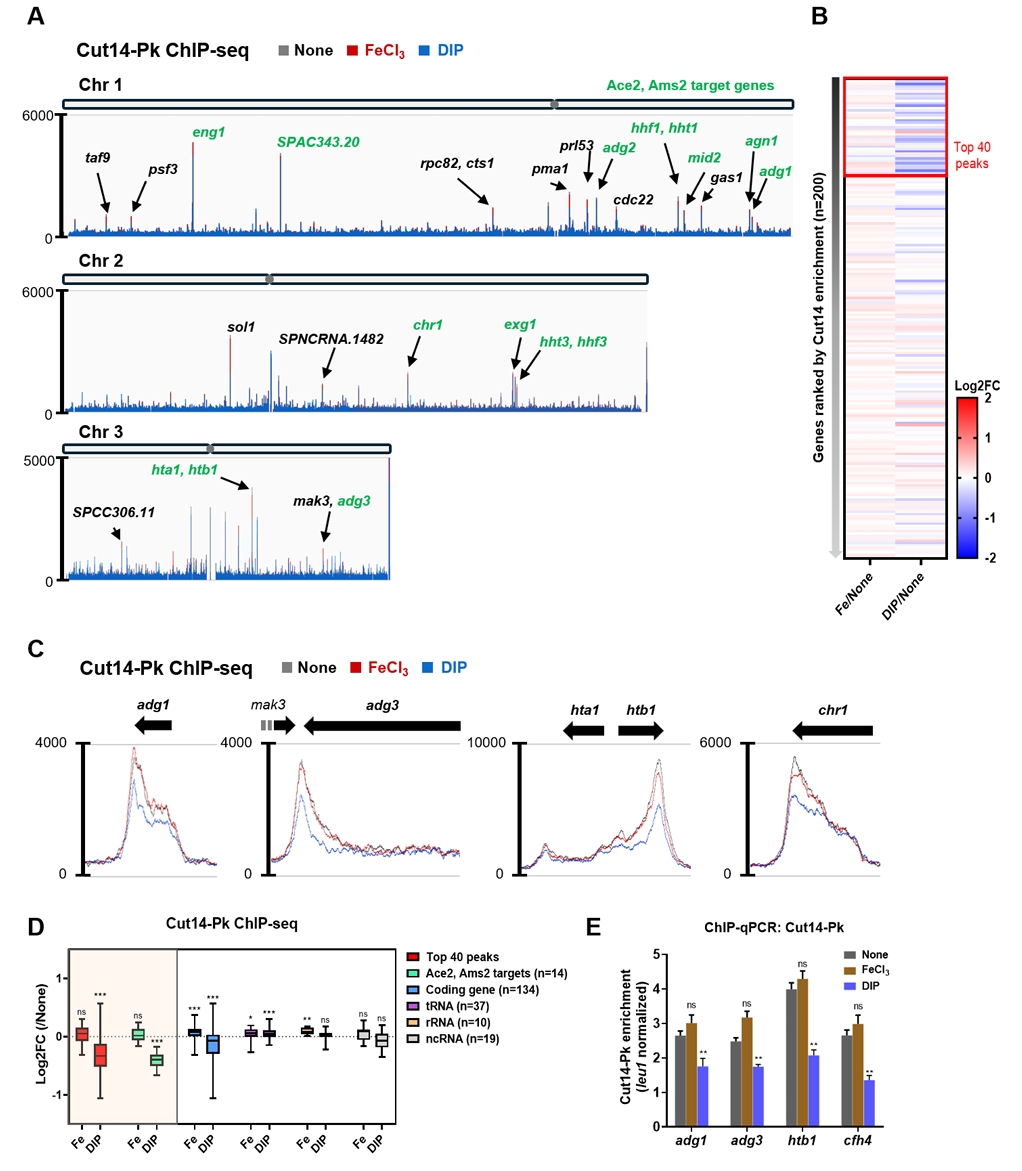
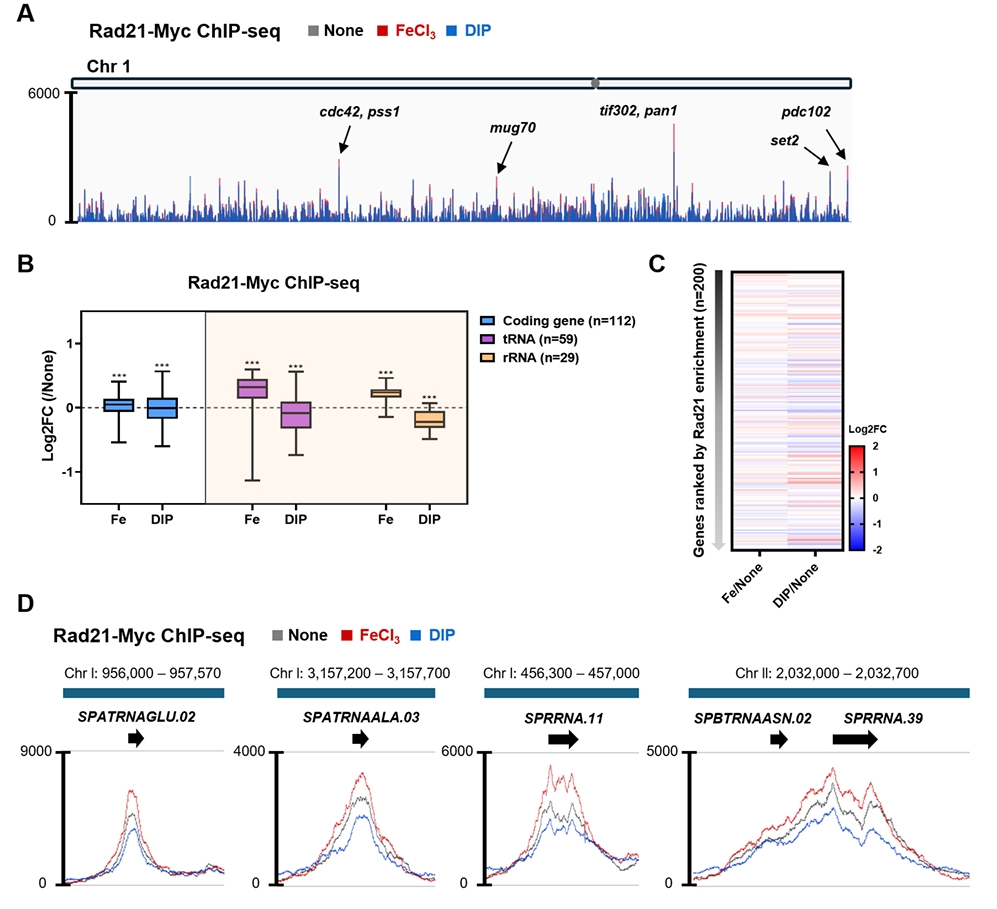
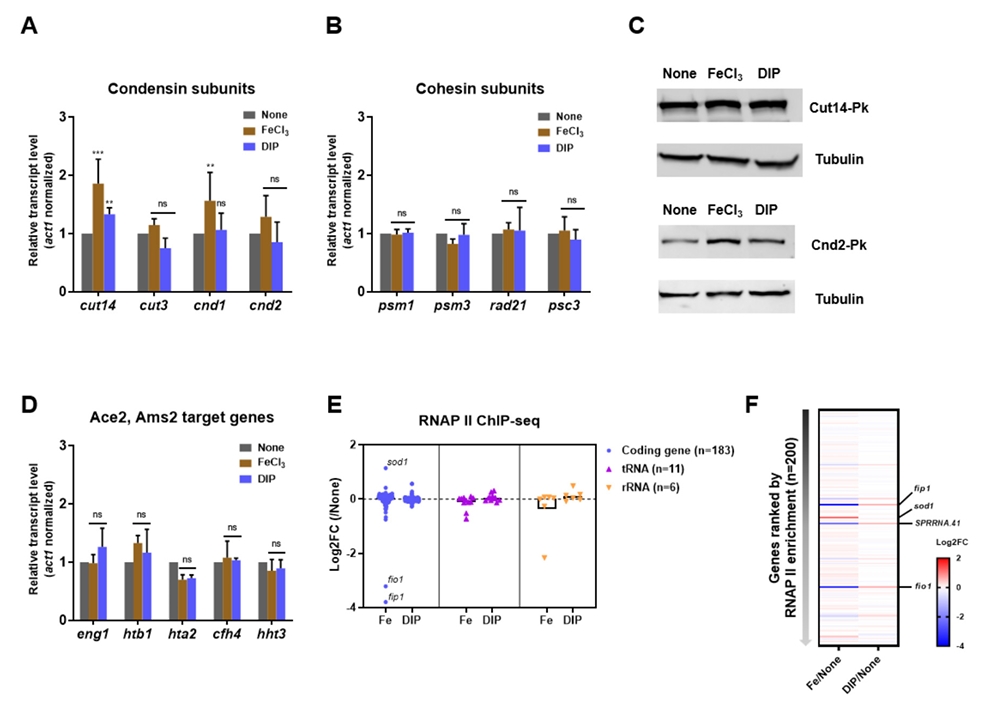
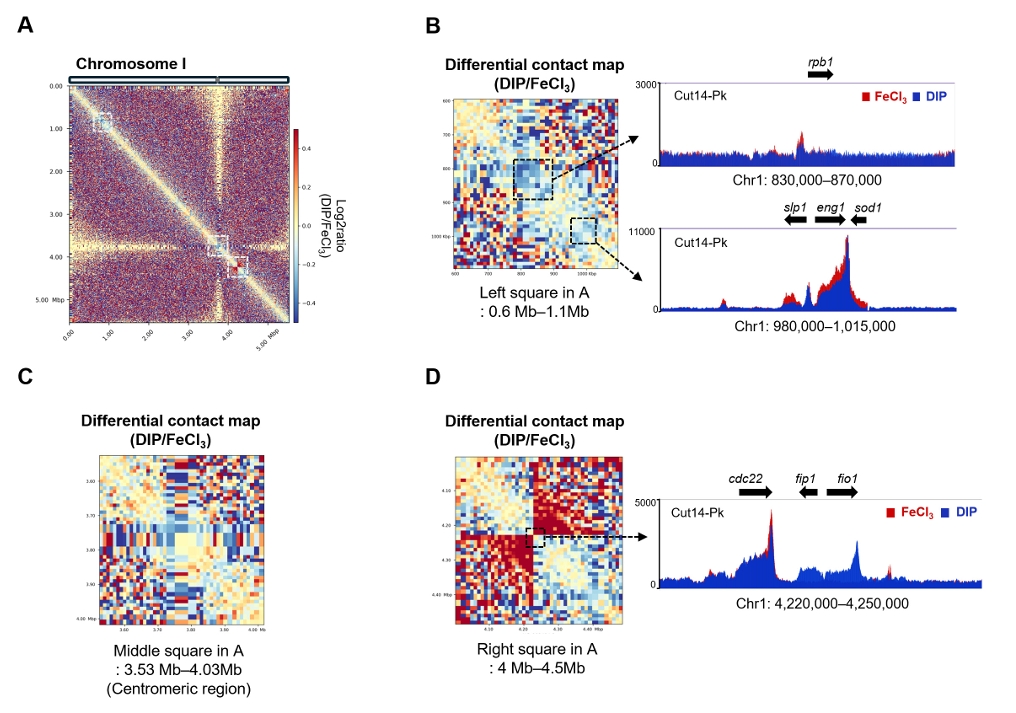
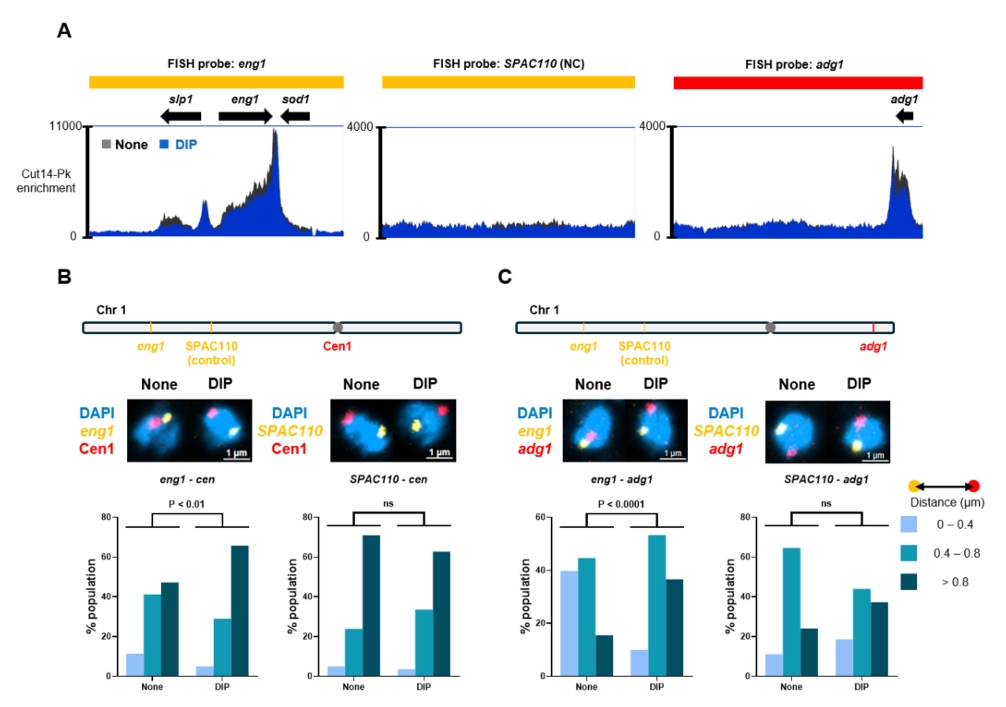
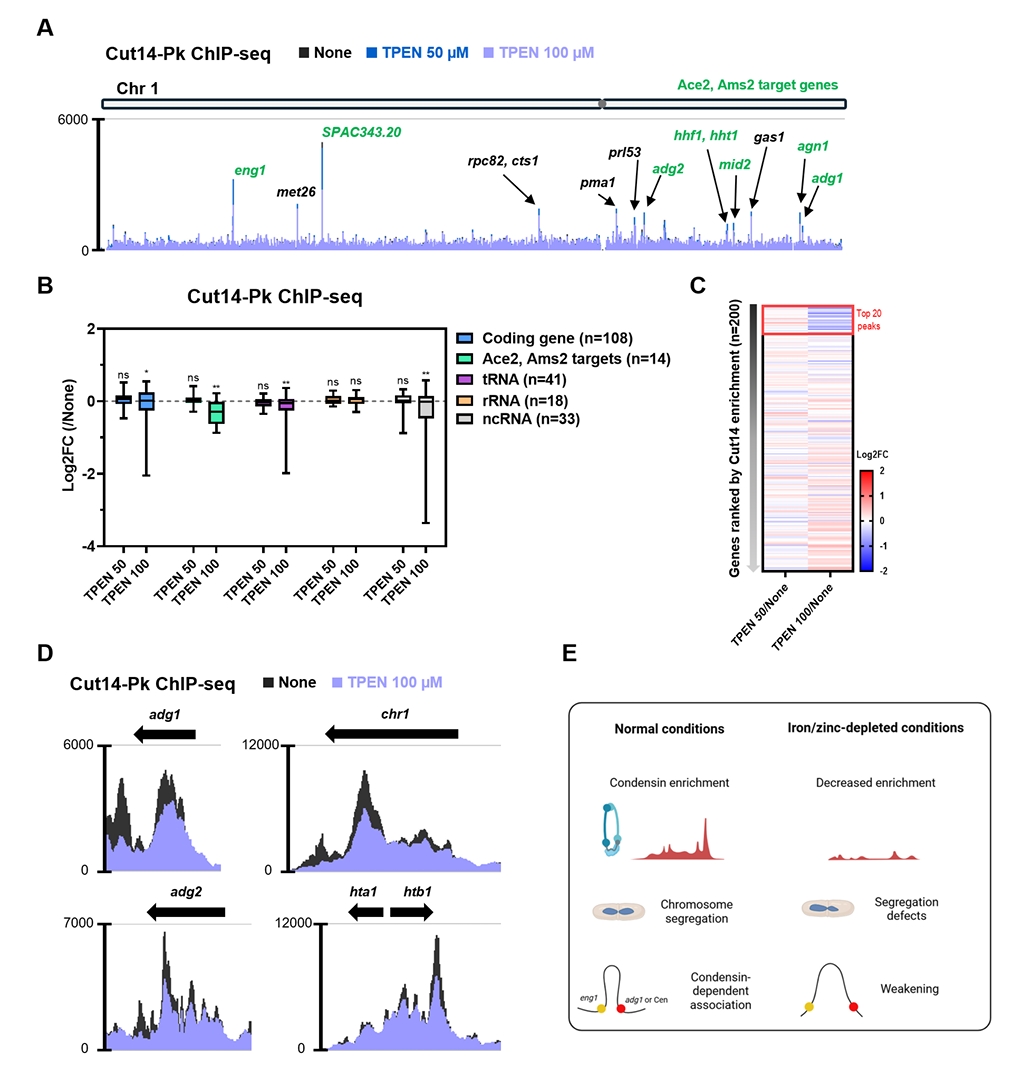
- Condensin plays a central role in mitotic chromosome organization and segregation by mediating long-range chromatin interactions. However, the extent to which cellular metabolic status influences condensin function remains unclear. To gain insights into the relationship of metal ion homeostasis and the function of condensin, we conducted genome-wide chromatin immunoprecipitation sequencing (ChIP-seq) using Schizosaccharomyces pombe under iron- or zinc-deficient conditions. Under iron- or zinc-deficient conditions, ChIP-seq results revealed a selective reduction in condensin binding at high-affinity target loci, particularly genes regulated by Ace2 and Ams2, while cohesin binding remained largely unaffected. Hi-C analysis showed that iron depletion weakened chromatin interactions at these condensin targets and centromeres, without disrupting global genome architecture. DNA fluorescence in situ hybridization (FISH) confirmed that iron deficiency impaired long-range associations between centromeres and Ace2 target loci at the single-cell level. Notably, iron deficiency led to chromosome segregation defects during mitosis, suggesting that diminished condensin occupancy compromised genome stability. These changes occurred without significant alterations in condensin protein levels or global transcription, indicating a direct effect of metal ion availability on condensin activity. Collectively, our findings revealed a previously unrecognized regulatory axis in which cellular metal ion homeostasis modulated condensin-dependent chromatin organization and mitotic chromosome segregation, offering new insights into the integration of metabolic state with genome maintenance.
Introduction
Materials and Methods
Results
Discussion
Acknowledgments
This work was supported by Chung-Ang University Research Grants (2023) and a grant from the National Research Foundation of Korea (NRF) awarded to K.K. (2022R1A2C1004423).
Conflict of Interest
The authors declare no conflict of interest.
Ethical Statements
Not applicable.
Supplementary Information
- Allshire RC, Nimmo ER, Ekwall K, Javerzat JP, Cranston G. 1995. Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes Dev. 9: 218–233. ArticlePubMed
- Bähler J, Wu JQ, Longtine MS, Shah NG, McKenzie III A, et al. 1998. Heterologous modules for efficient and versatile PCR‐based gene targeting in Schizosaccharomyces pombe. Yeast. 14: 943–951. ArticlePubMed
- Chikashige Y, Kinoshita N, Nakaseko Y, Matsumoto T, Murakami S, et al. 1989. Composite motifs and repeat symmetry in S. pombe centromeres: direct analysis by integration of NotI restriction sites. Cell. 57: 739–751. ArticlePubMed
- Chowdhary S, Kainth AS, Paracha S, Gross DS, Pincus D. 2022. Inducible transcriptional condensates drive 3D genome reorganization in the heat shock response. Mol Cell. 82: 4386–4399. ArticlePubMedPMC
- Corkins ME, May M, Ehrensberger KM, Hu Y-M, Liu Y-H, et al. 2013. Zinc finger protein Loz1 is required for zinc-responsive regulation of gene expression in fission yeast. Proc Natl Acad Sci USA. 110: 15371–15376. ArticlePubMedPMC
- Gadaleta MC, Iwasaki O, Noguchi C, Noma K-I, Noguchi E. 2015. Chromatin immunoprecipitation to detect DNA replication and repair factors. Methods Mol Biol. 1300: 169–186. ArticlePubMedPMC
- Gallagher PS, Larkin M, Thillainadesan G, Dhakshnamoorthy J, Balachandran V, et al. 2018. Iron homeostasis regulates facultative heterochromatin assembly in adaptive genome control. Nat Struct Mol Biol. 25: 372–383. ArticlePubMedPMCPDF
- Hirano T. 2012. Condensins: Universal organizers of chromosomes with diverse functions. Genes Dev. 26: 1659–1678. ArticlePubMedPMC
- Holtzen SE, Navid E, Kainov JD, Palmer AE. 2024. Transient Zn2+ deficiency induces replication stress and compromises daughter cell proliferation. Proc Natl Acad Sci USA. 121: e2321216121. ArticlePubMedPMC
- Iwasaki O, Tanaka A, Tanizawa H, Grewal SI, Noma KI. 2010. Centromeric localization of dispersed Pol III genes in fission yeast. Mol Biol Cell. 21: 254–265. ArticlePubMedPMC
- Kakui Y, Rabinowitz A, Barry DJ, Uhlmann F. 2017. Condensin-mediated remodeling of the mitotic chromatin landscape in fission yeast. Nat Genet. 49: 1553–1557. ArticlePubMedPMCPDF
- Killilea DW, Ames BN. 2008. Magnesium deficiency accelerates cellular senescence in cultured human fibroblasts. Proc Natl Acad Sci USA. 105: 5768–5773. ArticlePubMedPMC
- Kim K-D, Iwasaki O, Noma K. 2016a. An IF-FISH approach for covisualization of gene loci and nuclear architecture in fission yeast. Methods Enzymol. 574: 167–180. ArticlePubMedPMC
- Kim K-D, Tanizawa H, Iwasaki O, Noma K-i. 2016b. Transcription factors mediate condensin recruitment and global chromosomal organization in fission yeast. Nat Genet. 48: 1242–1252. ArticlePubMedPMCPDF
- Klug A. 2010. The discovery of zinc fingers and their applications in gene regulation and genome manipulation. Annu Rev Biochem. 79: 213–231. ArticlePubMed
- Labbé S, Pelletier B, Mercier A. 2007. Iron homeostasis in the fission yeast Schizosaccharomyces pombe. Biometals. 20: 523–537. ArticlePubMed
- Lengronne A, Katou Y, Mori S, Yokobayashi S, Kelly GP, et al. 2004. Cohesin relocation from sites of chromosomal loading to places of convergent transcription. Nature. 430: 573–578. ArticlePubMedPMCPDF
- Li L, Lyu X, Hou C, Takenaka N, Nguyen HQ, et al. 2015. Widespread rearrangement of 3D chromatin organization underlies polycomb-mediated stress-induced silencing. Mol Cell. 58: 216–231. ArticlePubMedPMC
- Mizuguchi T, Fudenberg G, Mehta S, Belton JM, Taneja N, et al. 2014. Cohesin-dependent globules and heterochromatin shape 3D genome architecture in S. Pombe. Nature. 516: 432–435. ArticlePubMedPMCPDF
- Muckenthaler MU, Rivella S, Hentze MW, Galy B. 2017. A red carpet for iron metabolism. Cell. 168: 344–361. ArticlePubMedPMC
- Nakazawa N, Arakawa O, Yanagida M. 2019. Condensin locates at transcriptional termination sites in mitosis, possibly releasing mitotic transcripts. Open Biol. 9: 190125.ArticlePubMedPMCLink
- Nasmyth K, Haering CH. 2009. Cohesin: Its roles and mechanisms. Annu Rev Genet. 43: 525–558. ArticlePubMed
- Niwa O. 2018. Determination of the frequency of minichromosome loss to assess chromosome stability in fission yeast. Cold Spring Harb Protoc. 2018: pdb.prot091991.ArticlePubMed
- Ocampo D, Damon LJ, Sanford L, Holtzen SE, Jones T, et al. 2024. Cellular zinc status alters chromatin accessibility and binding of p53 to DNA. Life Sci Alliance. 7: e202302327. ArticlePubMedPMC
- Ono T, Fang Y, Spector DL, Hirano T. 2004. Spatial and temporal regulation of condensins I and II in mitotic chromosome assembly in human cells. Mol Biol Cell. 15: 3296–3308. ArticlePubMedPMC
- Pelletier B, Trott A, Morano KA, Labbé S. 2005. Functional characterization of the iron-regulatory transcription factor Fep1 from Schizosaccharomyces pombe. J Biol Chem. 280: 25146–25161. ArticlePubMed
- Qin Y, Grimm SA, Roberts JD, Chrysovergis K, Wade PA. 2020. Alterations in promoter interaction landscape and transcriptional network underlying metabolic adaptation to diet. Nat Commun. 11: 962.ArticlePubMedPMCPDF
- Sissi C, Palumbo M. 2009. Effects of magnesium and related divalent metal ions in topoisomerase structure and function. Nucleic Acids Res. 37: 702–711. ArticlePubMedPMC
- Sutani T, Sakata T, Nakato R, Masuda K, Ishibashi M, et al. 2015. Condensin targets and reduces unwound DNA structures associated with transcription in mitotic chromosome condensation. Nat Commun. 6: 7815.ArticlePubMedPMCPDF
- Tanizawa H, Kim K-D, Iwasaki O, Noma K-i. 2017. Architectural alterations of the fission yeast genome during the cell cycle. Nat Struct Mol Biol. 24: 965–976. ArticlePubMedPMCPDF
- Wilson S, Liu YH, Cardona-Soto C, Wadhwa V, Foster MP, et al. 2019. The Loz1 transcription factor from Schizosaccharomyces pombe binds to Loz1 response elements and represses gene expression when zinc is in excess. Mol Microbiol. 112: 1701–1717. ArticlePubMedPMCLink
- Yan M, Song Y, Wong CP, Hardin K, Ho E. 2008. Zinc deficiency alters DNA damage response genes in normal human prostate epithelial cells. J Nutr. 138: 667–673. ArticlePubMed
- Zhang X-R, Zhao L, Suo F, Gao Y, Wu Q, et al. 2022. An improved auxin-inducible degron system for fission yeast. G3 (Bethesda). 12: jkab393.ArticlePubMedPMCPDF
References
Supplementary Information
References
Citations
 ePub Link
ePub Link Cite this Article
Cite this Article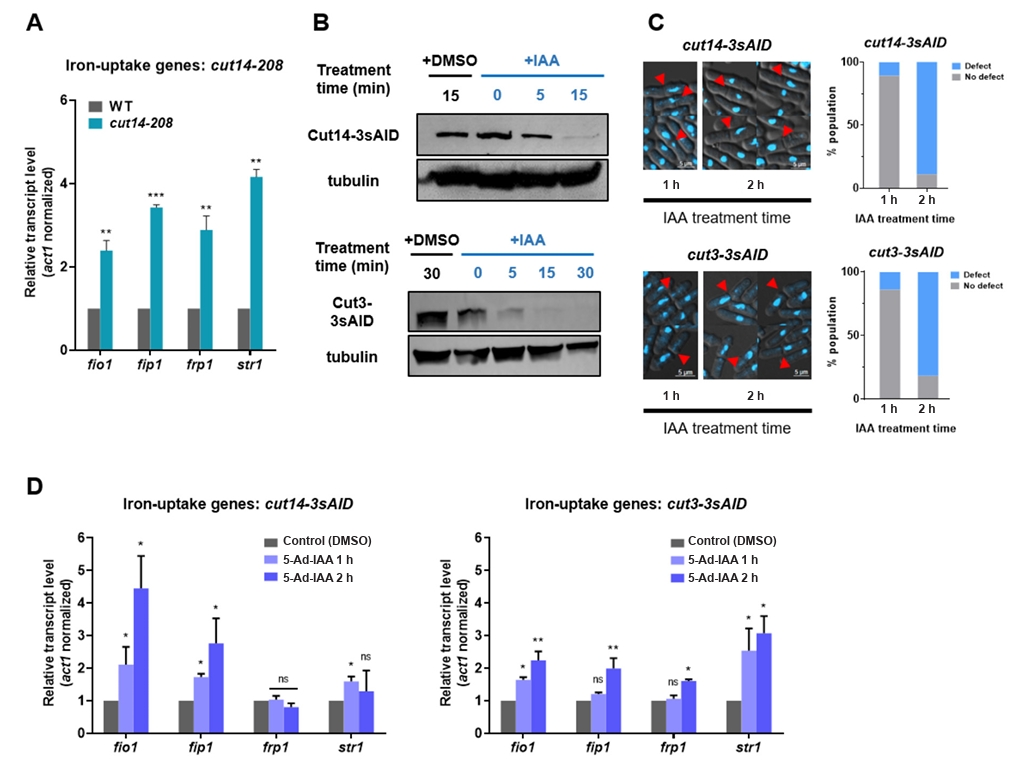
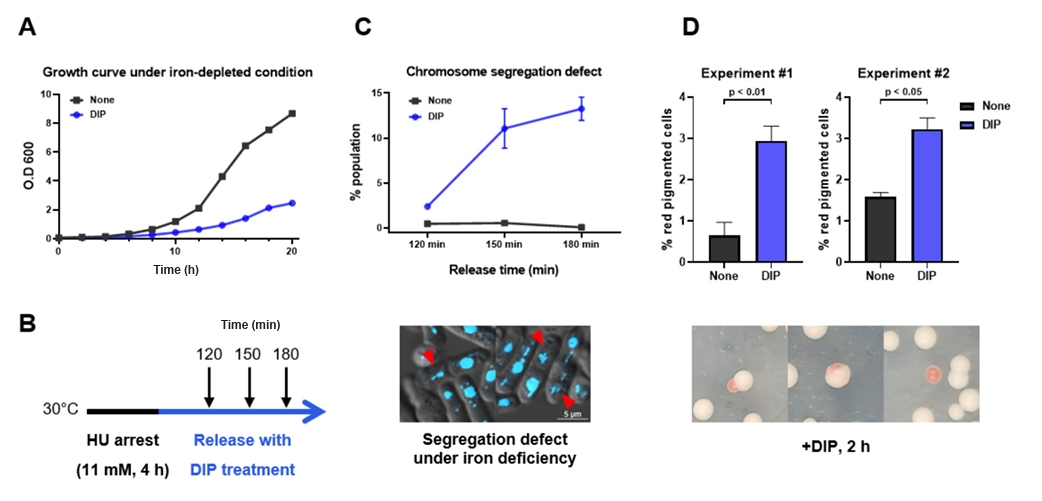
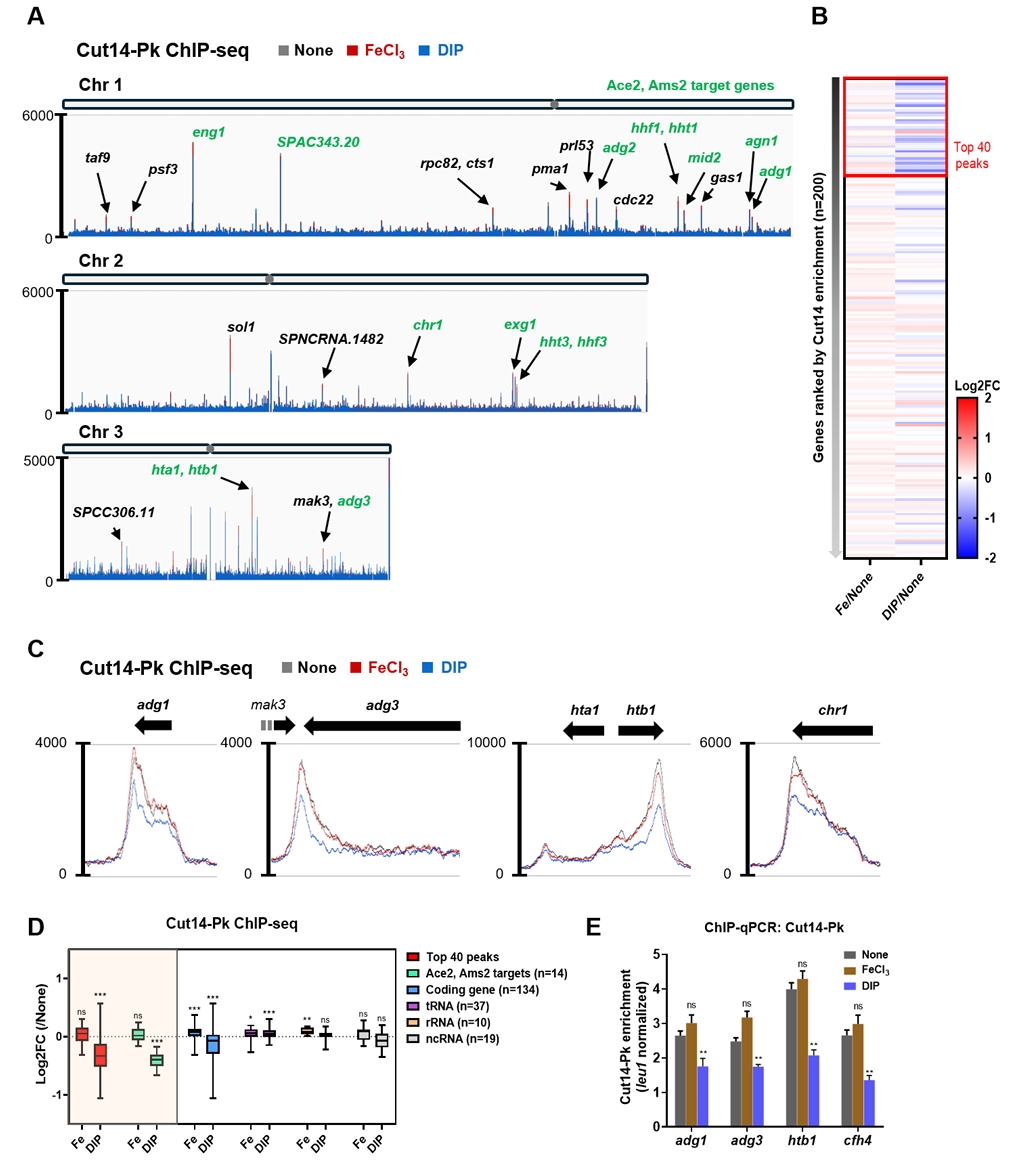
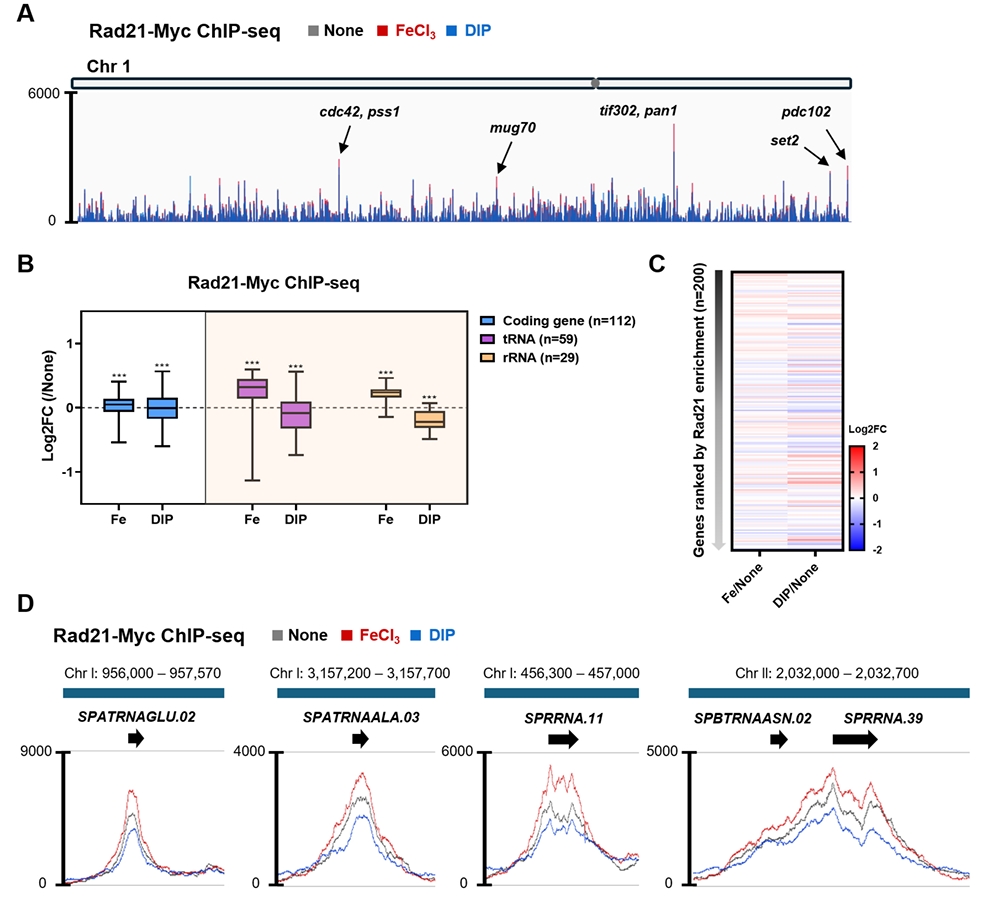
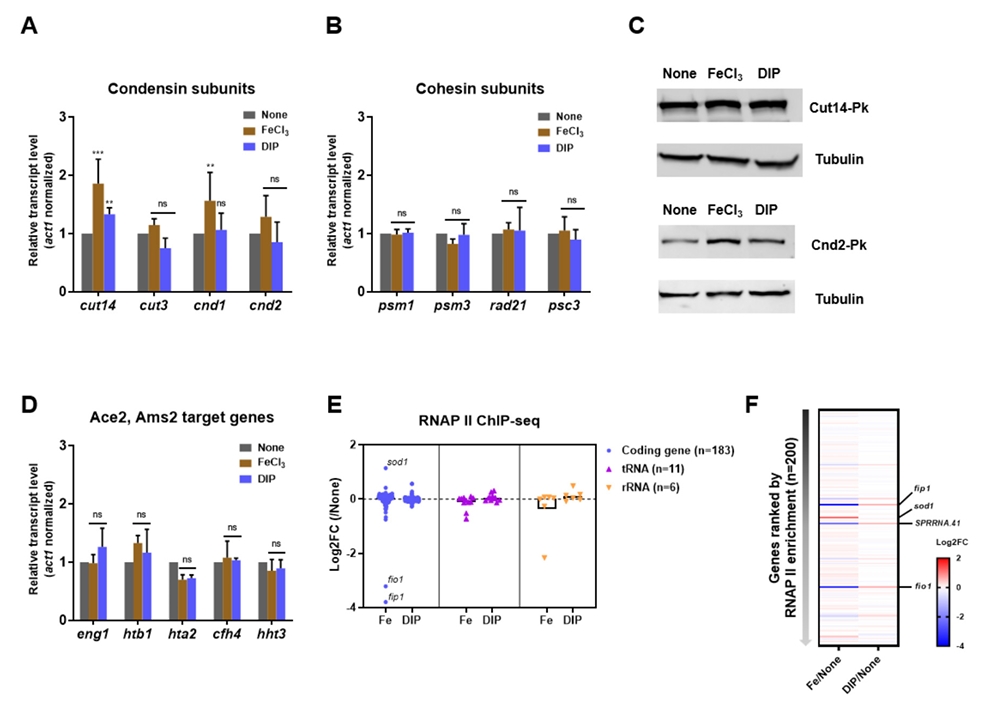
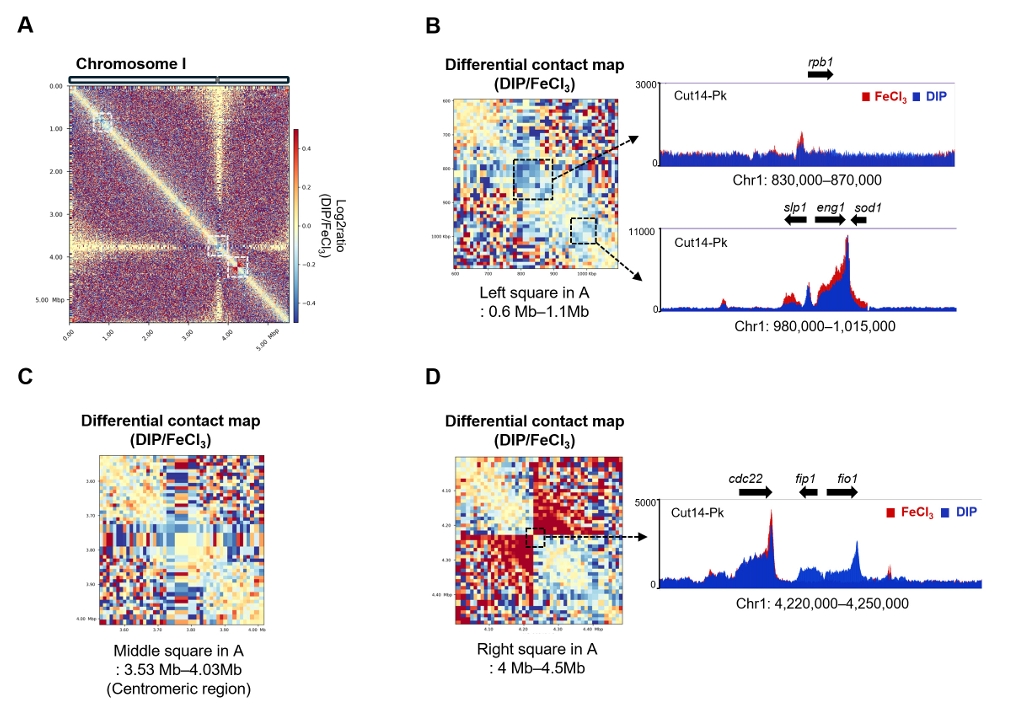
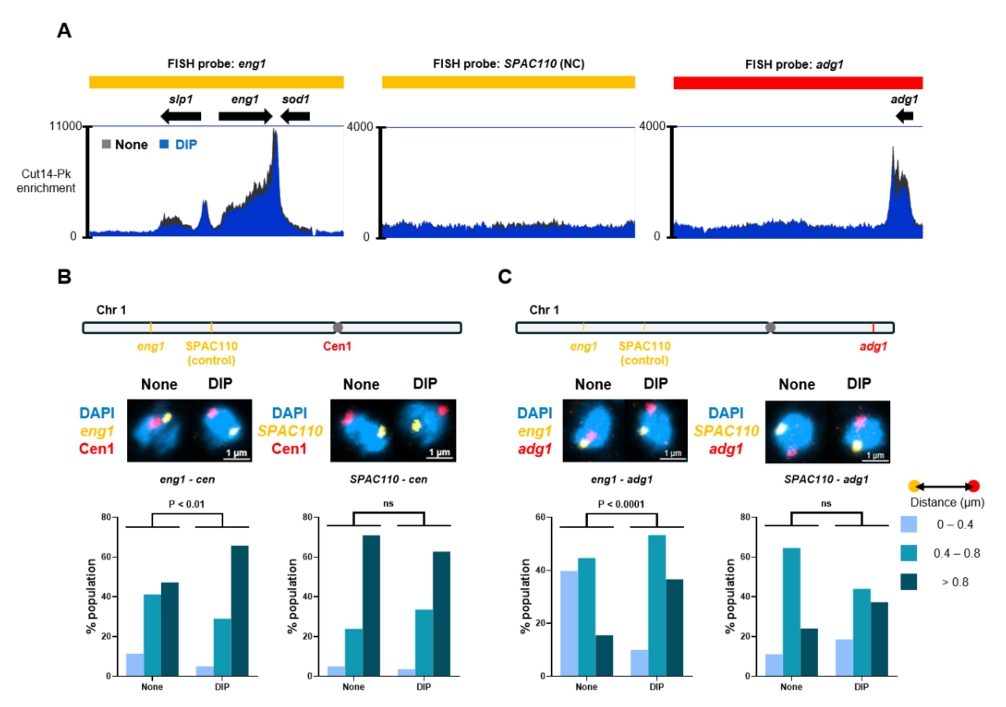
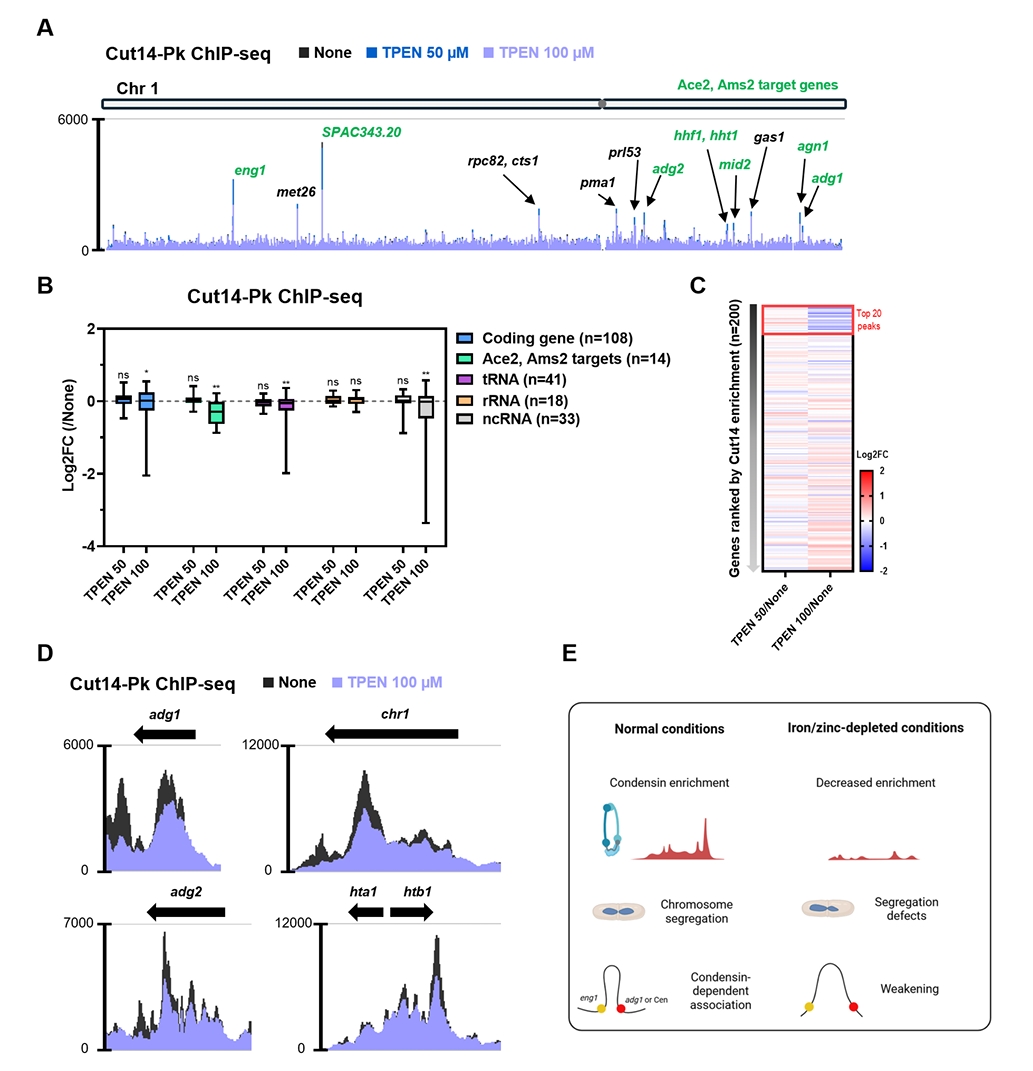








Fig. 1.
Fig. 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Fig. 8.
TOP

