ABSTRACT
- Basal cell carcinoma (BCC) is the most common form of skin cancer, with ultraviolet radiation recognized as the primary environmental driver; however, the potential contribution of alterations in the skin microbiota remains incompletely understood, particularly in Asian populations. This exploratory pilot study describes bacterial community patterns in BCC lesions compared with contralateral clinically normal skin in 20 Korean patients. Lesional and contralateral samples were obtained using paired skin swabs and punch biopsies and analyzed by full-length 16S rRNA gene sequencing, with targeted quantitative PCR (qPCR) of the roxP antioxidant gene of Cutibacterium acnes. Given the low-biomass nature of skin samples and the exploratory design, analyses focused on descriptive trends rather than confirmatory inference.
Across available samples, C. acnes was the dominant taxon, with a trend toward lower relative abundance in BCC lesions, particularly in biopsy-derived datasets. Microbial evenness appeared higher in lesions than controls. Predictive functional profiling suggested reduced representation of vitamin B6 metabolism pathways in lesions, while qPCR analysis of swab samples showed a trend toward lower roxP/16S rRNA ratios in BCC-associated microbiota. These findings should be interpreted cautiously in light of methodological constraints, including sample heterogeneity, lidocaine exposure prior to biopsy, absence of sequencing-based negative controls, and reliance on predictive functional inference.
Overall, this pilot study highlights potential differences in skin bacterial community structure between BCC lesions and contralateral skin in a Korean cohort. Larger, methodologically optimized studies incorporating metagenomic and functional validation will be required to determine whether these microbiota shifts contribute to, or result from, BCC-associated changes in the cutaneous environment.
-
Keywords: skin microbiota, basal cell carcinoma, Korean
Introduction
Basal cell carcinoma (BCC) is the most common malignancy of the skin and accounts for the majority of non-melanoma skin cancers worldwide. Although BCC rarely metastasizes, it can cause substantial local tissue destruction and morbidity. Ultraviolet radiation (UVR) is the primary environmental risk factor for BCC, inducing DNA damage, oxidative stress, and local immune dysregulation. However, BCC development is increasingly recognized as a multifactorial process influenced by host genetics, immune responses, and the local cutaneous microenvironment (Woo et al., 2022).
The skin microbiota plays an important role in maintaining cutaneous homeostasis through modulation of immune signaling, barrier integrity, and oxidative balance (Byrd et al., 2018; Woo and Kim, 2024). Alterations in bacterial community structure have been implicated in inflammatory skin disorders and, more recently, in skin carcinogenesis and UV-induced immune modulation (Nakatsuji et al., 2021; Patra et al., 2019). Experimental and clinical studies have demonstrated that UV exposure itself can perturb skin microbial composition and host–microbe interactions, supporting a mechanistic link between chronic photodamage and microbiota alteration (Burns et al., 2019; Grant et al., 2024; Patra et al., 2019).
In keratinocyte cancers and their precursors, emerging evidence suggests lesion-associated microbial shifts relative to surrounding skin. Microbiota differences have been reported in actinic keratosis and cutaneous squamous cell carcinoma, conditions that share cumulative UV exposure and field cancerization with BCC. These studies describe altered community composition, reduced dominance of sebaceous-site commensals, and increased microbial evenness in lesional or photodamaged skin, although findings vary across cohorts and methodologies (Krueger et al., 2022; Voigt et al., 2022; Wood et al., 2018). Despite growing interest in the role of the skin microbiota in non-melanoma skin cancers, most available data derive from Western populations, and studies specifically addressing BCC remain limited (Kim and Kim, 2019; Kim et al., 2023; Lee et al., 2019).
Among dominant skin commensals, Cutibacterium acnes has been proposed to contribute to oxidative homeostasis through multiple antioxidant systems, including the secreted protein RoxP. RoxP has been shown to neutralize extracellular reactive oxygen species in vitro, although its in vivo relevance relative to other bacterial antioxidant pathways remains incompletely defined (Allhorn et al., 2016; Cros et al., 2023). Whether changes in C. acnes abundance or antioxidant potential are associated with BCC remains unclear.
The present study was designed as an exploratory pilot investigation to describe bacterial community patterns in BCC lesions compared with contralateral clinically normal skin in Korean patients. Using paired swab and biopsy sampling with full-length 16S rRNA sequencing and targeted roxP quantification, we aimed to identify preliminary trends that may inform hypotheses for future mechanistic studies. Given the modest cohort size and methodological constraints inherent to low-biomass skin microbiota research, all findings are interpreted as descriptive rather than causal.
Materials and Methods
Study design and participants
This pilot study was conducted at the Department of Dermatology, Incheon St. Mary’s Hospital, The Catholic University of Korea, between July 2021 and December 2022. Twenty Korean patients with histologically confirmed BCC were enrolled after providing written informed consent (Table 1). Exclusion criteria included systemic antibiotic use within four weeks or topical antibiotic application within two weeks prior to sampling. The study was approved by the institutional ethics committee (OC21TISI0038).
Sampling procedures
For each participant, microbiota samples were collected from the BCC lesion and from contralateral clinically normal skin at an anatomically matched site whenever possible. Two sampling approaches were used: surface swabbing and punch biopsy.
Swab sampling was performed first using sterile flocked swabs pre-moistened with buffer (0.15 mol/L NaCl with 0.1% Tween 20), brushed 20 times across the target site in a crosswise direction. Punch biopsies (3 mm) were subsequently obtained after local anesthesia with 2% lidocaine; no antiseptic agents were applied prior to sampling. Lidocaine has documented antimicrobial activity and may influence biopsy-derived microbiota profiles; lesion and control sites were handled identically within each patient.
Samples were immediately placed in sterile tubes, stored at −80°C, and processed within six months. Prolonged frozen storage may affect DNA integrity, particularly for Gram-positive bacteria. Not all participants contributed both swab and biopsy samples, resulting in incomplete pairing for some analyses.
An independently processed air swab blank sequenced using a MiSeq V4–V5 workflow yielded 916 total reads. Detected taxa are provided for qualitative reference only and were not used for read subtraction or statistical decontamination due to differences in sequencing platform and amplicon region.
DNA extraction and sequencing
DNA was extracted using the DNeasy PowerSoil Pro Kit (Qiagen, Germany), which is optimized for environmental samples but may show reduced lysis efficiency for certain Gram-positive taxa, such as C. acnes. Negative extraction or PCR blanks were not sequenced for the PacBio dataset, precluding formal contaminant filtering.
Full-length 16S rRNA genes were amplified using primers 27F and 1492R and sequenced on the PacBio Sequel II platform. Bioinformatic processing was performed using QIIME2 with DADA2 for denoising and amplicon sequence variant (ASV) inference. Taxonomic assignment was conducted using the RDP database (trainset 18) (Bolyen et al., 2019; Callahan et al., 2016; Cole et al., 2009; Martin, 2011). Analyses were limited to species-level resolution.
Functional prediction
Predictive functional profiling was performed using PICRUSt2 to estimate KEGG pathway representation. Given the use of 16S rRNA amplicon data derived from low-biomass skin samples and incomplete reference genome representation of skin commensals, all functional predictions were considered exploratory and interpreted as inferred genomic potential rather than measured biological activity.
Quantification of roxP gene
Absolute quantification of roxP gene was performed by qPCR in swab samples with sufficient DNA yield (Andersson et al., 2019). Primer efficiencies and standard curves were validated prior to analysis, and roxP copy number was normalized to total bacterial 16S rRNA gene copy number. Each reaction was performed in technical duplicates. Primer specificity was validated in silico against RefSeq bacterial genomes and the NCBI nt database.
Statistical analysis
Statistical analyses were conducted in R (v4.1.2) and GraphPad Prism (v8.4.3). Paired statistical tests were used whenever complete lesion–control pairs were available; unpaired tests were applied only when pairing was incomplete. Alpha diversity metrics were compared using paired t-tests or Wilcoxon signed-rank tests as appropriate.
Beta diversity was assessed using PERMANOVA (adonis2, vegan), with subject ID included as a blocking factor when applicable. Because ANOSIM does not account for paired sampling, beta-diversity results are interpreted descriptively. PICRUSt-based functional predictions were compared using paired Wilcoxon signed-rank tests with Benjamini–Hochberg correction. Multivariate analyses, including Random Forest feature ranking and SparCC network inference, were considered exploratory and hypothesis-generating.
Results
Study cohort and sample overview
The cohort included 20 patients (12 female, 8 male; mean age 79.0 years, range 53–100). Most lesions were located on the face, head, or neck, with one perianal lesion (Table 1). Comorbidities such as diabetes mellitus, hypertension, chronic kidney disease, and dyslipidemia were common and are acknowledged as potential confounders.
A total of 15 biopsy pairs and 13 swab pairs were available for analysis.
Microbial composition and diversity
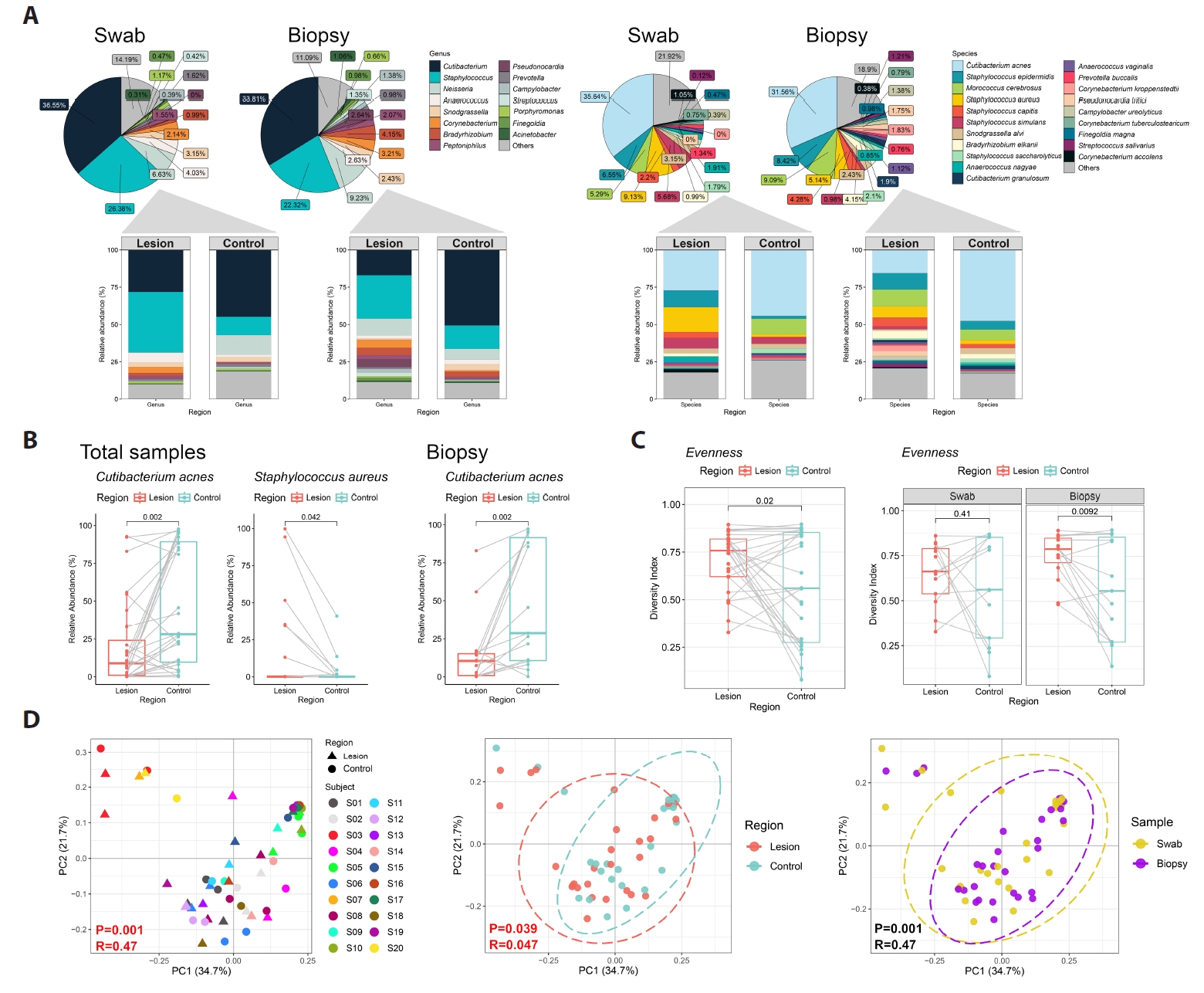
Across all samples, sequencing identified 450 bacterial species spanning 243 genera, 28 classes, and 13 phyla. The dominant genera were Cutibacterium and Staphylococcus (Figs. 1A and 2A). At the species level, C. acnes was the most abundant taxon. In biopsy-derived datasets, C. acnes showed a trend toward lower relative abundance in BCC lesions compared with contralateral skin, while swab datasets showed a similar but less pronounced pattern (Fig. 1B).
Microbial evenness tended to be higher in lesions, whereas richness showed no consistent difference (Fig. 1C).
Exploratory multivariate analyses
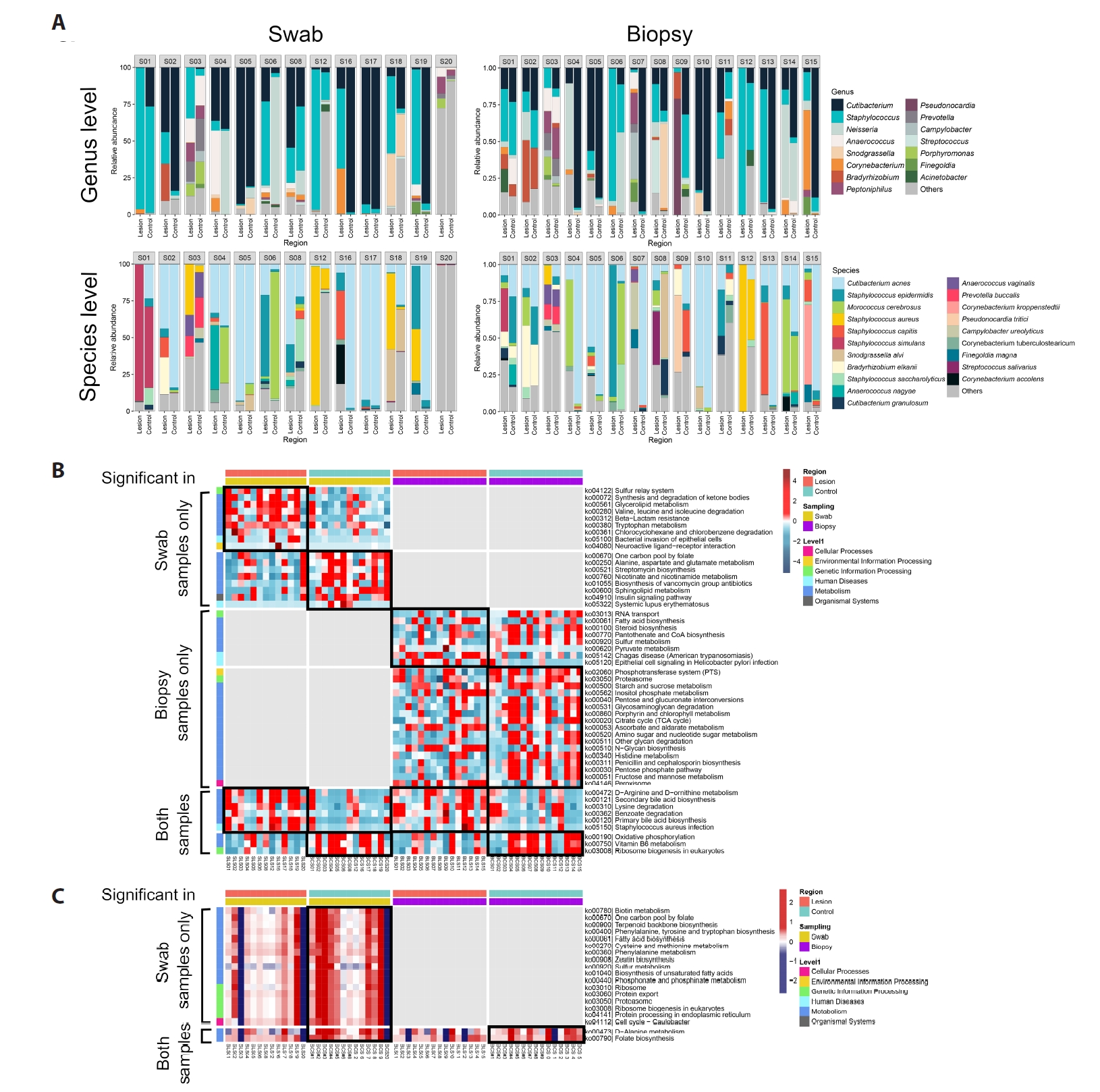
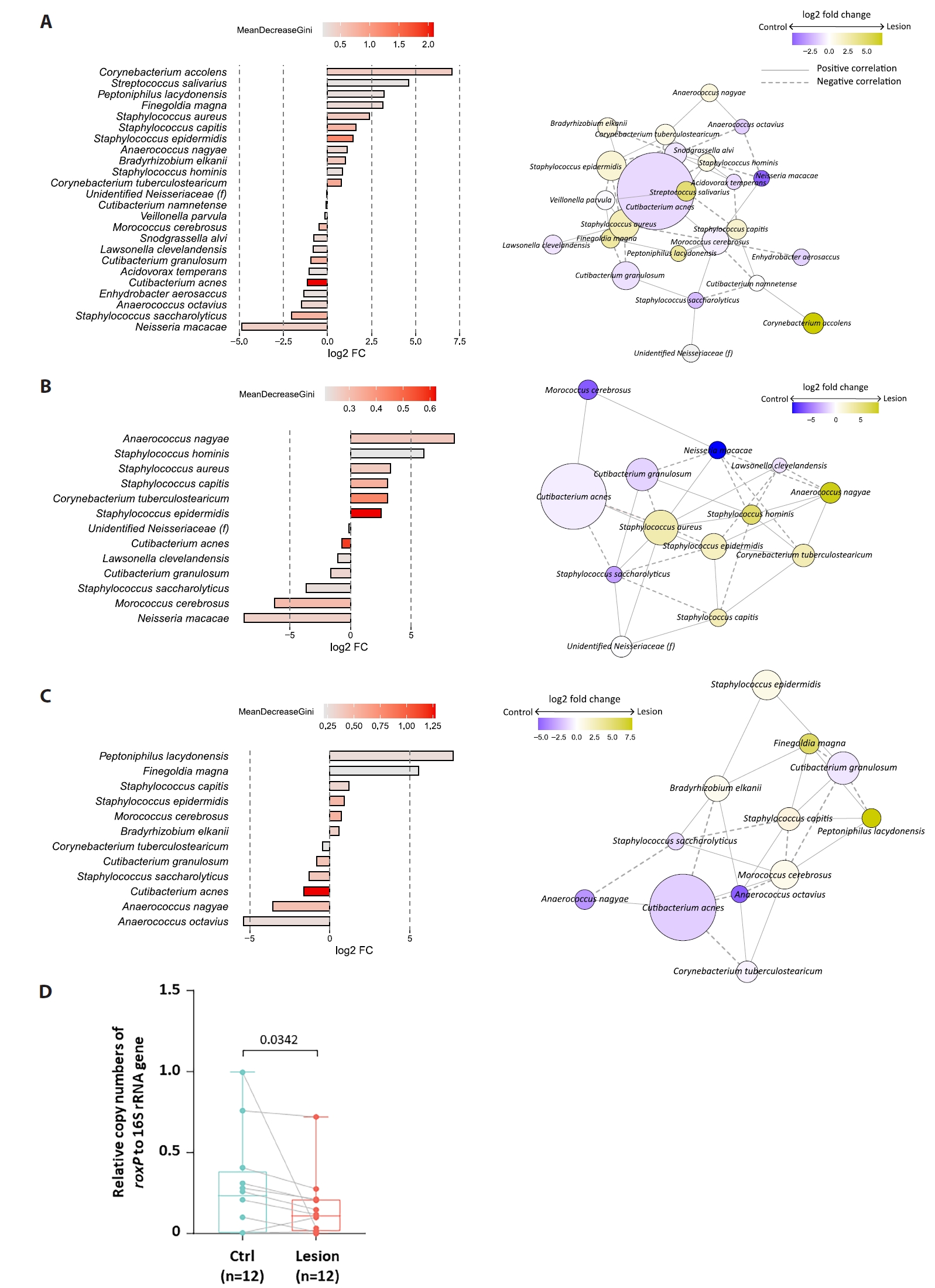
Beta diversity analyses showed partial clustering by individual, with modest separation between lesion and control samples (Fig. 1D). Random Forest analysis ranked C. acnes among the top taxa contributing to lesion–control differentiation (Fig. 3A). For swab samples, the model exhibited modest discrimination (AUC = 0.71) with low out-of-bag accuracy, whereas biopsy-based models demonstrated near-random performance (AUC = 0.56); accordingly, given the small sample size and high dimensionality, Random Forest analyses were treated as exploratory and interpreted as descriptive, serving primarily for pattern identification and feature ranking rather than predictive inference. SparCC network analysis suggested altered interspecies associations in lesion samples (Fig. 3A–3C), but these patterns require validation.
Predicted functional profiles
PICRUSt2-based inference suggested lower representation of vitamin B6 metabolism pathways in BCC lesions and relatively higher folate biosynthesis in controls, particularly among C. acnes-associated ASVs (Fig. 2B–2C). These predictions are exploratory and hypothesis-generating.
roxP gene quantification
In 12 paired swab samples, the roxP/16S ratio tended to be lower in BCC lesions than in controls, with substantial inter-individual variability (Fig. 3D). Given the limited sample size and potential confounding by overall C. acnes abundance, these results are interpreted cautiously.
Discussion
This exploratory pilot study describes differences in skin bacterial community structure between BCC lesions and contralateral clinically normal skin in a Korean cohort. The most consistent trend was a relative reduction of C. acnes in lesion samples, particularly in biopsy-derived datasets, accompanied by increased microbial evenness. Similar community-level features—namely reduced dominance of sebaceous-associated taxa and increased evenness—have been reported in photodamaged skin and in keratinocyte cancers such as actinic keratosis and cutaneous squamous cell carcinoma (Burns et al., 2019; Grant et al., 2024; Voigt et al., 2022), although direct comparisons are limited by methodological differences.
Previous studies in predominantly Western populations have reported that chronic UV exposure and non-melanoma skin cancers are associated with shifts in cutaneous microbial communities, including reduced dominance of sebaceous-site commensals and increased community evenness or diversity in lesional or photodamaged skin (Patra et al., 2019; Woo et al., 2022). Microbiota studies in actinic keratosis and SCC further suggest that microbial composition may vary across the spectrum of UV-induced field cancerization (Krueger et al., 2022; Voigt et al., 2022; Wood et al., 2018), supporting the plausibility that BCC-associated lesions may also exhibit distinct microbial patterns. UV radiation has been shown to directly alter bacterial viability and metabolism, as well as indirectly shape microbial communities through effects on skin barrier function, immune responses, and oxidative stress (Nakatsuji et al., 2021; Patra et al., 2019). In this context, the observed reduction of C. acnes in BCC lesions in our study aligns with prior observations that sebaceous-associated taxa may be depleted in environments characterized by chronic inflammation, tissue remodeling, or tumor-associated microenvironmental changes. Importantly, data on skin microbiota in Asian patients with BCC remain scarce, and our findings extend existing literature by providing preliminary insights from a Korean cohort.
Methodological limitations substantially constrain interpretation. Biopsy sampling required lidocaine anesthesia, which may suppress bacterial viability. DNA extraction bias, prolonged frozen storage, low microbial biomass, and the absence of sequencing-based negative controls further limit quantitative inference. Anatomical site heterogeneity and systemic comorbidities may also influence observed microbiota patterns.
The observed reduction in roxP abundance likely reflects decreased C. acnes representation rather than reduced antioxidant capacity per bacterium. C. acnes encodes multiple oxidative stress defense systems, including catalases, peroxidases, and thioredoxin-related pathways, and roxP represents only one component of its broader redox biology. While RoxP has been shown to exert extracellular antioxidant activity in vitro (Andersson et al., 2019; Brüggemann et al., 2021), its relative contribution to oxidative balance on human skin in vivo remains incompletely defined. Accordingly, roxP should be viewed as a surrogate marker of community shifts rather than a dominant functional determinant.
Predictive functional differences observed in this study should be interpreted with particular caution. PICRUSt2 results represent inferred genomic potential rather than direct evidence of in vivo activity. Although reference genomes indicate that C. acnes harbors genes involved in folate and one-carbon metabolism, 16S-based inference cannot determine whether these pathways are transcriptionally active or biologically relevant within the BCC microenvironment. Future studies incorporating shotgun metagenomics and functional assays will be required.
Conclusion
This pilot study provides preliminary evidence of altered skin microbiota composition in BCC lesions compared with contralateral skin in Korean patients. While trends suggest reduced C. acnes representation and altered inferred functional potential, substantial methodological limitations preclude causal inference. Larger, methodologically optimized studies are required to clarify the role of skin microbiota in BCC-associated microenvironmental changes.
Abbreviations
Basal cell carcinoma, BCC; Ultraviolet radiation, UVR; Control, Ctrl; Phylogenetic investigation of communities by reconstruction of unobserved states, PICRUSt2; Random forest, RF; Analysis of similarities, ANOSIM; Sparse correlations for compositional data, SparCC; Radical oxygenase of Propionibacterium acnes, RoxP; Quantitative polymerase chain reaction, qPCR; Principal coordinate analysis, PCoA; Amplicon sequence variant, ASV.
Acknowledgments
This work was supported by a Grant of Translational R&D Project through the Institute for Bio-Medical convergence, Incheon St. Mary’s Hospital, The Catholic University of Korea, the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (grant number: 2023R1A2C1007759) and a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number: RS-2023-KH-136575 and RS-2025-02217860).
Author Contributions
Conceptualization: HSK; Data curation: HSK, SK; Formal analysis: HLK, IC, AK; Funding Acquisition: HSK; Investigation: HSK, SK; Methodology: HSK, HLK, IC, AK; Project administration: HSK; Resources: HLK, AK, WJS; Software: AK, WJS; Supervision: HSK; Writing–Original Draft Preparation: HSK; Writing–Review and Editing: AK, WJS.
Conflict of Interest
The authors declare no conflict of interest.
Data Availability
Sequencing data will be deposited in the NCBI Sequence Read Archive (SRA) prior to publication; accession number(s) will be provided in the revised proof.
Ethics Statement
The study was approved by the Ethics Committee of Incheon St. Mary’s Hospital, The Catholic University of Korea (approval number OC21TISI0038). All participants provided their written informed consent to publication of their case details. The authors thank patients with BCC for their contributions.
Supplementary Information
The online version contains supplementary material available at https://doi.org/10.71150/jm.2511012.
Fig. 1.Skin microbiota profiling in basal cell carcinoma (BCC) among Korean patients (n = 20). (A) Relative abundance of the most prevalent bacterial genera and species in paired swab (n = 13 pairs) and biopsy (n = 15 pairs) samples. (B) Bacterial species showing differences in mean relative abundance between BCC lesions and contralateral clinically normal skin (Control, Ctrl) in the combined dataset (swab and biopsy) and the biopsy-only dataset. (C) Alpha diversity (Evenness) comparisons between BCC lesions and controls across the total dataset, biopsy dataset, and swab dataset. (D) Principal coordinates analysis (PCoA) based on weighted UniFrac distances, visualized by individual, sampling method, and skin site.
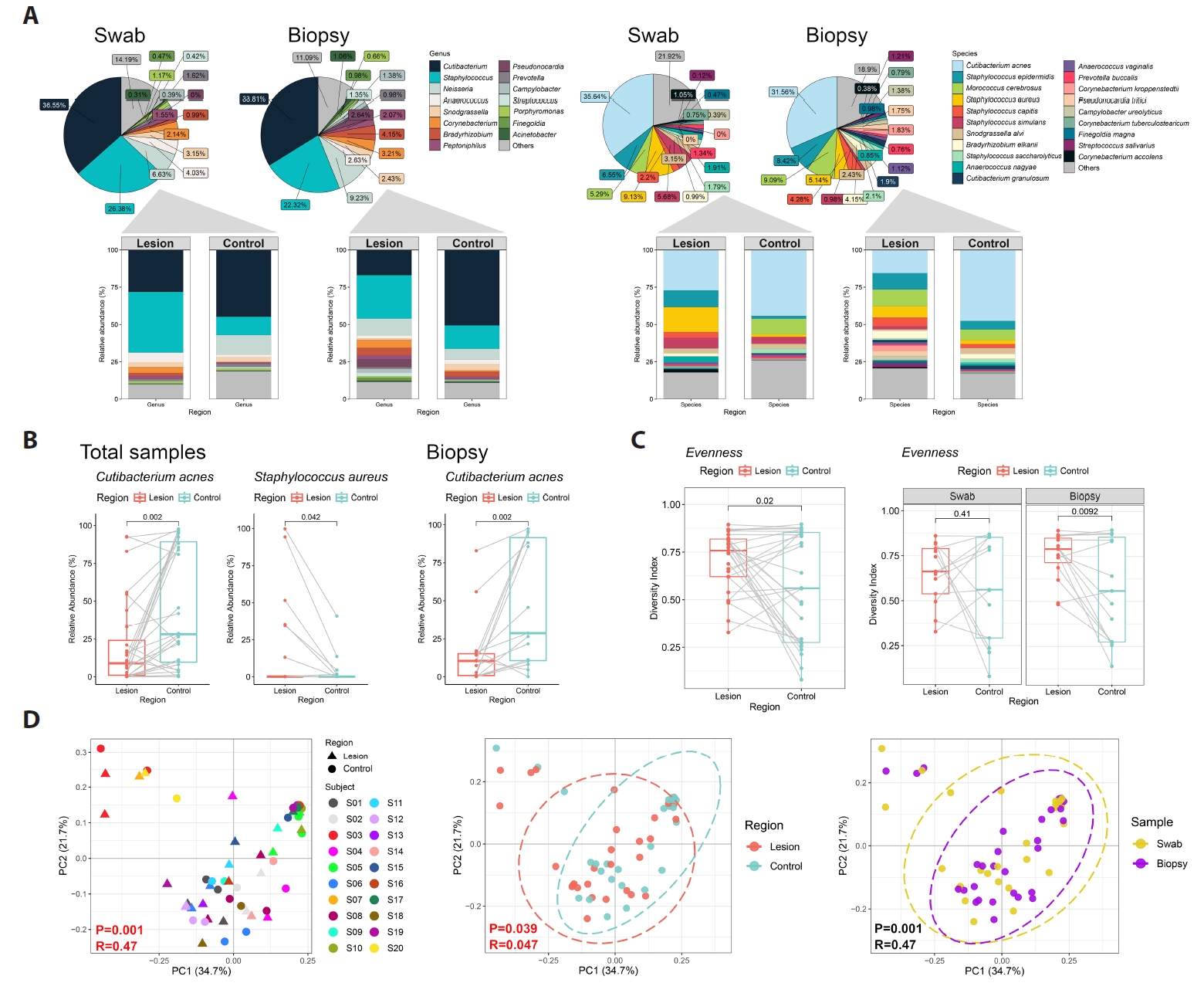
Fig. 2.Microbiota composition and predicted functional profiles in BCC. (A) Relative abundance of the most prevalent bacterial genera and species in swab (n = 13 pairs) and biopsy (n = 15 pairs) datasets. (B) Predicted metagenomic functional pathways (PICRUSt2) showing the top differentially abundant KEGG pathways between BCC lesions and controls. (C) Comparison of C. acnes–associated KEGG pathway proportions between BCC lesions and controls.
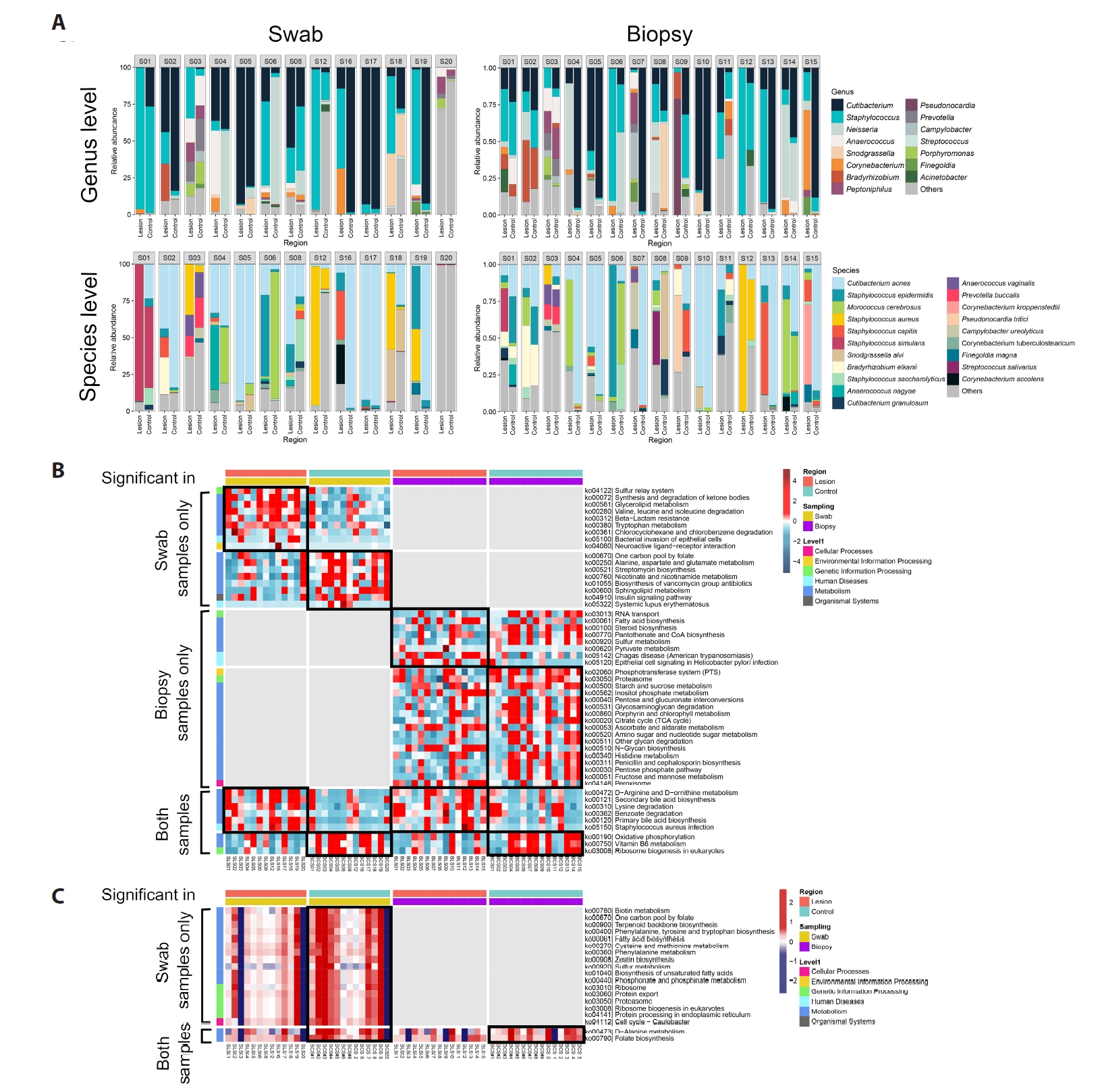
Fig. 3.Discriminatory taxa and roxP antioxidant gene quantification in BCC. (A–C) Random forest analysis identifying the top 24 taxa (mean decrease Gini ≥ 0.2) distinguishing BCC lesions from controls in the total dataset (A), swab dataset (B), and biopsy dataset (C). Right panels: co-occurrence networks of amplicon sequence variants (ASVs) selected by random forest. Edges represent significant SparCC correlations (P < 0.05), with solid lines indicating positive and dashed lines indicating negative associations. Node size and color represent log₂ fold change between BCC and control. (D) Absolute quantification of the roxP gene (Radical oxygenase of Propionibacterium acnes) in swab samples (n = 12 pairs), expressed as the ratio of roxP to 16S rRNA copy number. Differences assessed by paired Wilcoxon signed-rank test; P < 0.05.
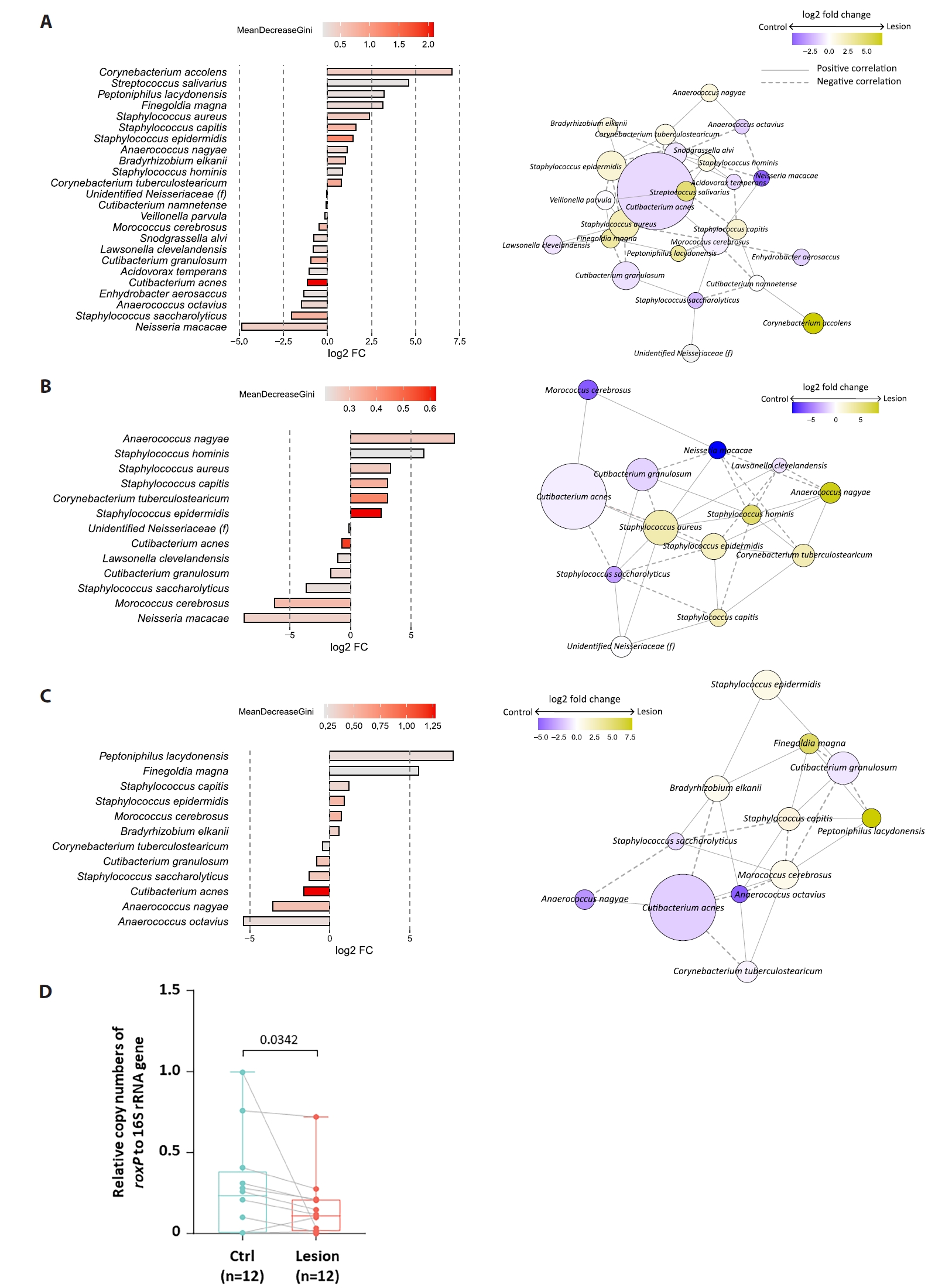
Table 1.Patient characteristics
|
Subject number |
Sex/Age |
Race |
Diagnosis |
Pathology |
Pigment |
Location |
Swab |
Biopsy |
Co-morbidity |
|
1 |
F/77 |
Korean |
BCC |
Nodulocystic |
Present |
Back |
O |
O |
Peripheral neural disease |
|
2 |
M/87 |
Korean |
BCC |
Superficial |
Present |
Back |
O |
O |
DM |
|
3 |
F/100 |
Korean |
BCC |
Nodulocystic |
Present |
Cheek |
O |
O |
HTN |
|
4 |
M/68 |
Korean |
BCC |
Nodulocystic |
Present |
Nose |
O |
O |
HTN |
|
5 |
M/53 |
Korean |
BCC |
Superficial |
Present |
Nose |
O |
O |
None |
|
6 |
F/82 |
Korean |
BCC |
Micronodular |
Present |
Cheek |
O |
O |
Vitiligo |
|
7 |
M/71 |
Korean |
BCC |
Nodulocystic |
Present |
Perinasal area |
X |
O |
None |
|
8 |
M/73 |
Korean |
BCC |
Nodulocystic |
Present |
Lower eyelid |
O |
O |
DM, dyslipidemia |
|
9 |
F/86 |
Korean |
BCC |
Nodulocystic |
Present |
Cheek |
X |
O |
HTN |
|
10 |
M/71 |
Korean |
BCC |
Nodulocystic |
Present |
Nose |
X |
O |
Parkinson’s disease, cataract, HTN, DM |
|
11 |
F/68 |
Korean |
BCC |
Superficial |
Present |
Back |
X |
O |
HTN |
|
12 |
F/80 |
Korean |
BCC |
Nodulocystic |
Present |
Upper eyelid |
O |
O |
Stroke |
|
13 |
M/81 |
Korean |
BCC |
Nodulocystic |
Present |
Nose |
X |
O |
DM, HTN, CKD |
|
14 |
M/62 |
Korean |
BCC |
Superficial |
Present |
Perioral area |
X |
O |
None |
|
15 |
M/82 |
Korean |
BCC |
Nodulocystic |
Present |
Perinasal area |
X |
O |
HTN |
|
16 |
F/97 |
Korean |
BCC |
Superficial |
Present |
Perioral area |
O |
X |
DM, HTN, CKD |
|
17 |
M/74 |
Korean |
BCC |
Micronodular |
Present |
Cheek |
O |
X |
HTN |
|
18 |
F/74 |
Korean |
BCC |
Nodulocystic |
Absent |
Nose |
O |
X |
DM |
|
19 |
F/79 |
Korean |
BCC |
Nodulocystic |
Present |
Lower eyelid |
O |
X |
HTN, dyslipidemia |
|
20 |
F/80 |
Korean |
BCC |
Superficial |
Present |
Perianal area |
O |
X |
None |
References
- Allhorn M, Arve S, Brüggemann H, Lood R. 2016. A novel enzyme with antioxidant capacity produced by the ubiquitous skin colonizer Cutibacterium acnes. Sci Rep. 6: 36412.ArticlePubMedPMC
- Andersson T, Ertürk Bergdahl G, Saleh K, Magnúsdóttir H, Stødkilde K, et al. 2019. Common skin bacteria protect their host from oxidative stress through secreted antioxidant RoxP. Sci Rep. 9: 3596.ArticlePubMedPMCPDF
- Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet CC, et al. 2019. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol. 37: 852–857. ArticlePubMedPMC
- Brüggemann H, Salar-Vidal L, Gollnick HPM, Lood R. 2021. A Janus-faced bacterium: host-beneficial and -detrimental roles of Cutibacterium acnes. Front Microbiol. 12: 673845.ArticlePubMedPMC
- Burns EM, Ahmed H, Isedeh PN, Kohli I, Van Der Pol W, et al. 2019. Ultraviolet radiation, both UVA and UVB, influences the composition of the skin microbiome. Exp Dermatol. 28: 136–141. ArticlePubMedPMC
- Byrd AL, Belkaid Y, Segre JA. 2018. The human skin microbiome. Nat Rev Microbiol. 16: 143–155. ArticlePubMedPDF
- Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJ, et al. 2016. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods. 13: 581–583. ArticlePubMedPMCPDF
- Cole JR, Wang Q, Cardenas E, Fish J, Chai B, et al. 2009. The ribosomal database project: improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 37: D141–D145. ArticlePubMed
- Cros MP, Mir-Pedrol J, Toloza L, Knödlseder N, Maruotti J, et al. 2023. New insights into the role of Cutibacterium acnes-derived extracellular vesicles in inflammatory skin disorders. Sci Rep. 13: 16058.ArticlePubMedPMC
- Grant GJ, Kohli I, Mohammad TF. 2024. A narrative review of the impact of ultraviolet radiation and sunscreen on the skin microbiome. Photodermatol Photoimmunol Photomed. 40: e12943. ArticlePubMed
- Kim HS, Keum HL, Chung IY, Nattkemper L, Head CR, et al. 2023. Characterization of a perturbed skin microbiome in prurigo nodularis and lichen simplex chronicus. J Invest Dermatol. 143: 2082–2085. ArticlePubMed
- Kim JE, Kim HS. 2019. Microbiome of the skin and gut in atopic dermatitis (AD): understanding the pathophysiology and finding novel management strategies. J Clin Med. 8: 444.ArticlePubMedPMC
- Krueger A, Zaugg J, Lachner N, Bialasiewicz S, Lin LL, et al. 2022. Changes in the skin microbiome associated with squamous cell carcinoma in transplant recipients. ISME Commun. 2: 13.ArticlePubMedPMCPDF
- Lee YB, Byun EJ, Kim HS. 2019. Potential role of the microbiome in acne: a comprehensive review. J Clin Med. 8: 987.ArticlePubMedPMC
- Martin M. 2011. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 17: 3.Article
- Nakatsuji T, Cheng JY, Gallo RL. 2021. Mechanisms for control of skin immune function by the microbiome. Curr Opin Immunol. 72: 324–330. ArticlePubMed
- Patra V, Wagner K, Arulampalam V, Wolf P. 2019. Skin microbiome modulates the effect of ultraviolet radiation on cellular response and immune function. iScience. 15: 211–222. ArticlePubMedPMC
- Voigt AY, Emiola A, Johnson JS, Fleming ES, Nguyen H, et al. 2022. Skin microbiome variation with cancer progression in human cutaneous squamous cell carcinoma. J Invest Dermatol. 142: 2773–2782. ArticlePubMedPMC
- Woo YR, Cho SH, Lee JD, Kim HS. 2022. The human microbiota and skin cancer. Int J Mol Sci. 23: 1813.ArticlePubMedPMC
- Woo YR, Kim HS. 2024. Interaction between the microbiota and the skin barrier in aging skin: a comprehensive review. Front Physiol. 15: 1322205.ArticlePubMedPMC
- Wood DLA, Lachner N, Tan JM, Tang S, Angel N, et al. 2018. A natural history of actinic keratosis and cutaneous squamous cell carcinoma microbiomes. mBio. 9: e01432-18.ArticlePubMedPMCLink
Citations
Citations to this article as recorded by

- Skin Microbiome Patterns Associated with Basal Cell Carcinoma: A Case Series
Mavra Masood, David Ozog, Tengfei Ma, Marissa Ceresnie, Aunna Pourang, Christine C. Johnson, Xinyue Qiu, Albert Levin, Jesse Veenstra
Microorganisms.2026; 14(4): 822. CrossRef


 ePub Link
ePub Link Cite this Article
Cite this Article