- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- For Contributors
- Policies
- E-Submission

- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- Policies
- For Contributors
Articles
- Page Path
- HOME > J. Microbiol > Volume 64(5); 2026 > Article
-
Full article
Delineated domain of VP2 capsid protein in H-1 parvovirus that determines susceptibility to human cancer cells - Il-Rae Cho1,†, Patcharporn Budluang1,†,#, Yeon Ha Kim2, Haan Park2, Namuk Kim2, Kon Ho Lee3, Jin-Hyun Ahn4, Ho Young Kang2,*, Young-Hwa Chung1,*
-
Journal of Microbiology 2026;64(5):e2601003.
DOI: https://doi.org/10.71150/jm.2601003
Published online: May 27, 2026
1Department of Cogno-Mechatronics Engineering, Optomechatronics Research Institute, Pusan National University, Busan 46241, Republic of Korea
2Department of Integrated Biological Science, Pusan National University, Busan 46241, Republic of Korea
3Department of Microbiology, School of Medicine, Gyeongsang National University, Jinju 52828, Republic of Korea
4Department of Microbiology, Sungkyunkwan University School of Medicine, Suwon 16419, Republic of Korea
- *Correspondence Ho Young Kang hoykang@pusan.ac.kr Young-Hwa Chung younghc@pusan.ac.kr
- †These authors contributed equally to this work.
#Present address: Department of Pharmacology, Faculty of Medicine, Chiang Mai University, Chiang Mai 50200, Thailand
© The Microbiological Society of Korea
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- 33 Views
- 5 Download
ABSTRACT
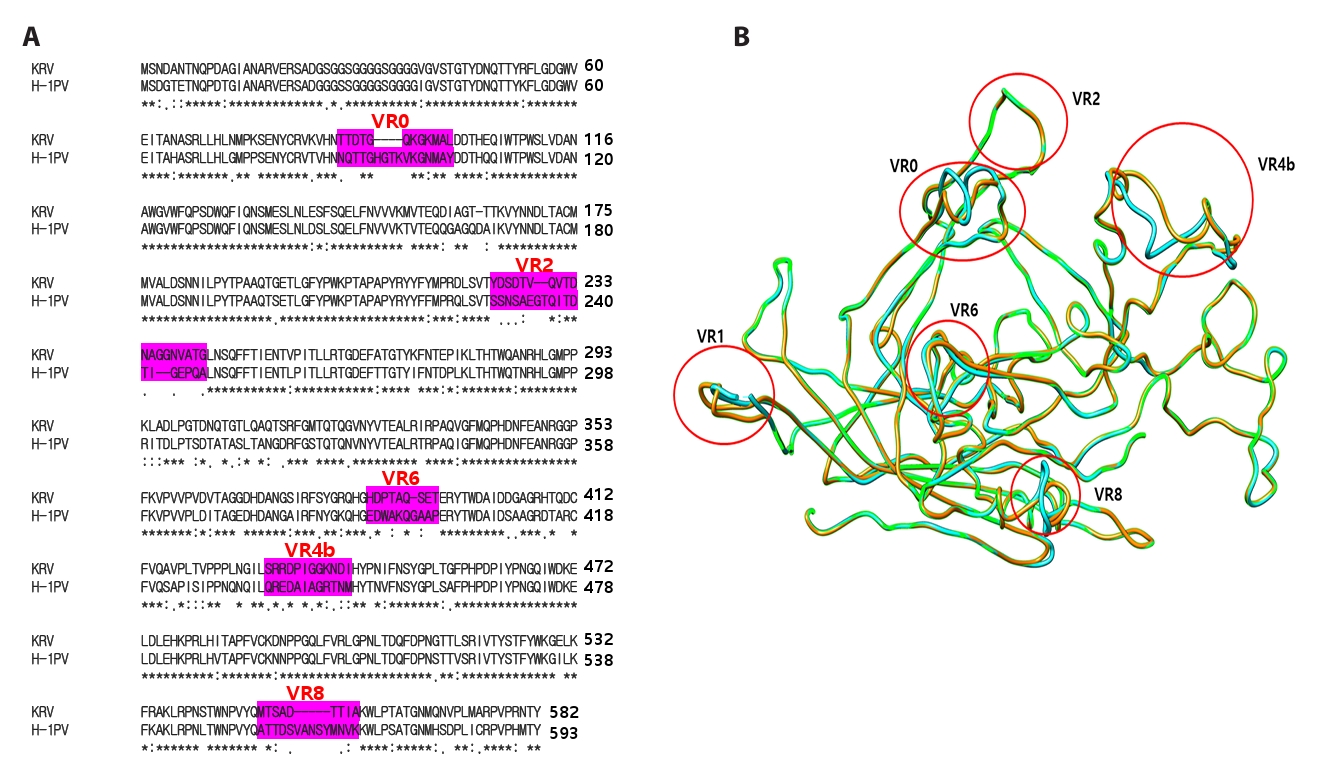
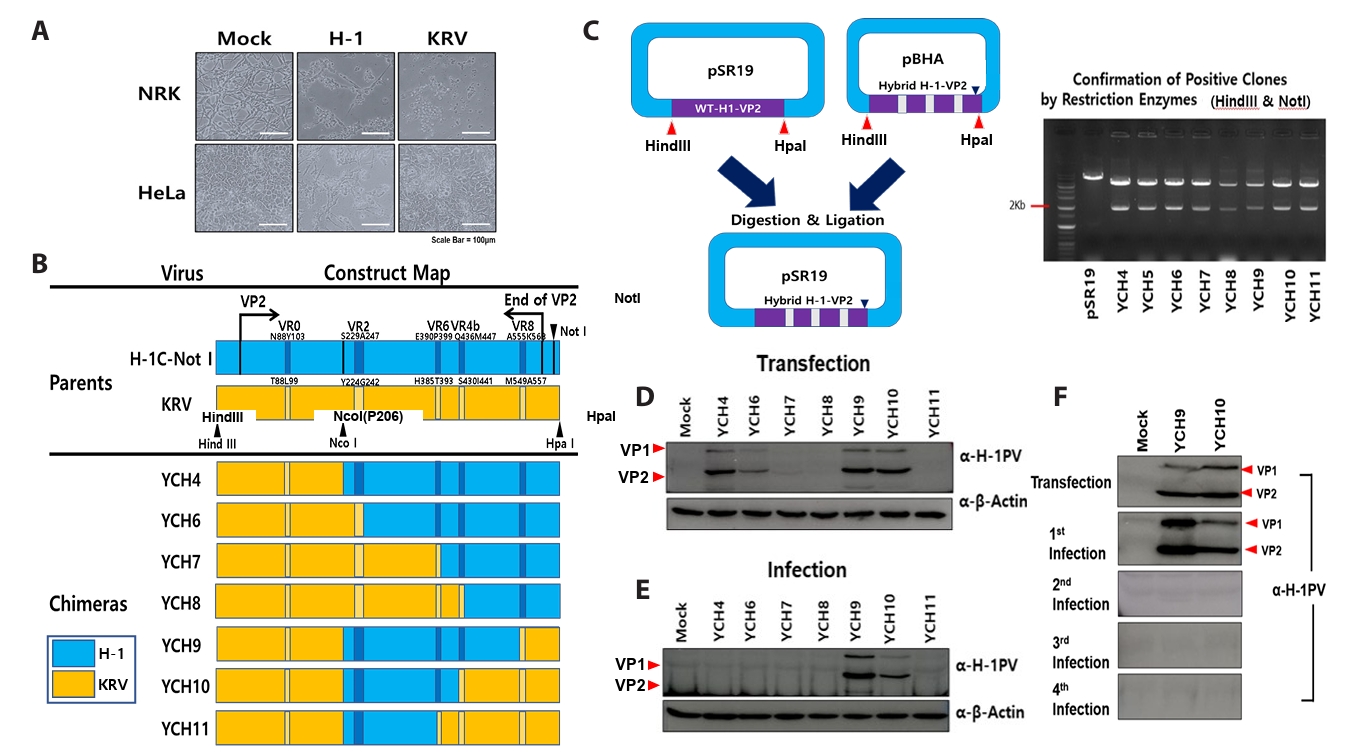
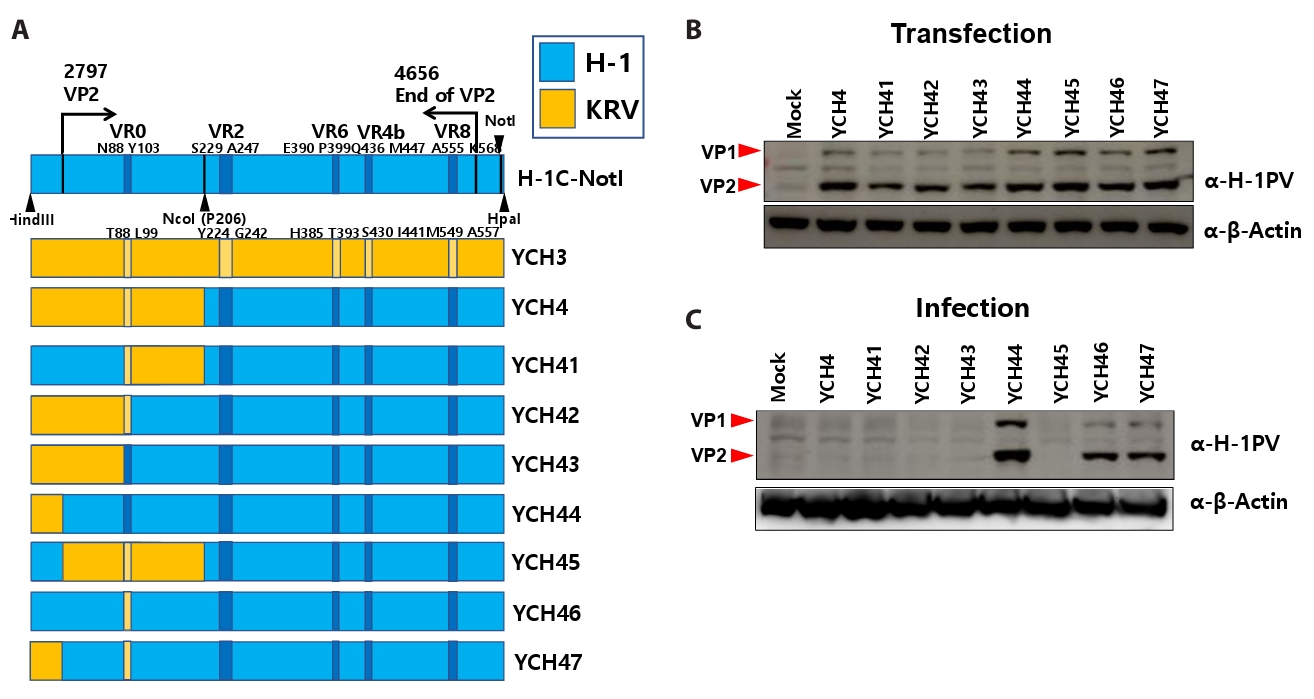
- Despite the application of H-1 parvovirus as an anticancer drug, the relationship between its specific tropism and oncolytic activity has been unknown. H-1 viral infection induced cytopathic effects in HeLa cells, whereas Kilham rat virus (KRV), similar to H-1 virus, did not. To explore which segments of the viral protein 2 (VP2) capsid protein in the H-1 virus determine susceptibility to human cancer cells, chimeric H-1 viruses with specific gene segments of H-1 VP2 were constructed. Delineation of the VP2 capsid protein revealed a minimum domain (K208–L435 in the H-1 VP2 protein) to determine infectivity in human cancer cells; however, this domain was not sufficient to maintain infectivity. To solve this problem, further construction of chimeric H-1 viruses illustrated the necessity of segments covering both M1-N87 and D104-P206 in the H-1 VP2 protein, based on chimeric H-1 viruses designated as YCH44, YCH45, and YCH46. Both the variable region 4b (VR4b) domains from KRV VP2 and VR8 from H-1 VP2 were required for the same purpose, based on chimeric H-1 viruses designated as YCH-HK8, YCH16, YCH17, YCH18, and YCH19. We confirmed that chimeric viruses carrying these segments infected human lung adenocarcinoma A549 and pancreatic cancer Panc-1 cells, whereas the parental KRV did not. Taken together, these findings indicate that specific domains of the H-1 virus VP2 capsid protein determine infectivity toward human cancer cells.
Introduction
Materials and Methods
Results
Discussion
Acknowledgments
This study was supported by a two-year research grant from Pusan National University, Republic of Korea.
Conflict of Interest
The authors declare no competing interests.
Supplementary Information
Supplementary Data S1.
- Allaume X, El-Andaloussi N, Leuchs B, Bonifati S, Kulkarni A, et al. 2012. Retargeting of rat parvovirus H-1PV to cancer cells through genetic engineering of the viral capsid. J Virol. 86: 3452–3465. ArticlePubMedPMCLink
- Ball-Goodrich LJ, Tattersall P. 1992. Two amino acid substitutions within the capsid are coordinately required for acquisition of fibrotropism by the lymphotropic strain of minute virus of mice. J Virol. 66: 3415–3423. ArticlePubMedPMCLink
- Beljanski V, Hiscott J. 2012. The use of oncolytic viruses to overcome lung cancer drug resistance. Curr Opin Virol. 2: 629–635. ArticlePubMed
- Besselsen DG, Besch-Williford CL, Pintel DJ, Franklin CL, Hook RR Jr, et al. 1995. Detection of H-1 parvovirus and Kilham rat virus by PCR. J Clin Microbiol. 33: 1699–1703. ArticlePubMedPMCLink
- Christensen J, Tattersall P. 2002. Parvovirus initiator protein NS1 and RPA coordinate replication fork progression in a reconstituted DNA replication system. J Virol. 76: 6518–6531. ArticlePubMedPMCLink
- Chung YH, Jun HS, Son M, Bao M, Bae HY, et al. 2000. Cellular and molecular mechanism for Kilham rat virus-induced autoimmune diabetes in DR-BB rats. J Immunol. 165: 2866–2876. ArticlePubMedPDF
- Conry RM, Westbrook B, Mckee S, Norwood TG. 2018. Talimogene laherparepvec: first in class oncolytic virotherapy. Hum Vaccin Immunother. 14: 839–846. ArticlePubMedPMC
- Di Piazza M, Mader C, Geletneky K, Herrero YCM, Weber E, et al. 2007. Cytosolic activation of cathepsins mediates parvovirus H-1-induced killing of cisplatin and TRAIL-resistant glioma cells. J Virol. 81: 4186–4198. ArticlePubMedPMCLink
- Fakhiri J, Schneider MA, Puschhof J, Stanifer M, Schildgen V, et al. 2019. Novel chimeric gene therapy vectors based on adeno-associated virus and four different mammalian bocaviruses. Mol Ther Methods Clin Dev. 12: 202–222. ArticlePubMedPMC
- Geiss C, Kis Z, Leuchs B, Frank-Stohr M, Schlehofer JR, et al. 2017. Preclinical testing of an oncolytic parvovirus: standard protoparvovirus H-1PV efficiently induces osteosarcoma cell lysis in vitro. Viruses. 9: 301.ArticlePubMedPMC
- Guberski DL, Thomas VA, Shek WR, Like AA, Handler ES, et al. 1991. Induction of type I diabetes by Kilham's rat virus in diabetes-resistant BB/Wor rats. Science. 254: 1010–1013. ArticlePubMed
- Gunther M, May P. 1976. Isolation and structural characterization of monomeric and dimeric forms of replicative intermediates of Kilham rat virus DNA. J Virol. 20: 86–95. ArticlePubMedPMCLink
- Hajda J, Leuchs B, Angelova AL, Frehtman V, Rommelaere J, et al. 2021. Phase 2 trial of oncolytic H-1 parvovirus therapy shows safety and signs of immune system activation in patients with metastatic pancreatic ductal adenocarcinoma. Clin Cancer Res. 27: 5546–5556. ArticlePubMedPDF
- Halder S, Nam HJ, Govindasamy L, Vogel M, Dinsart C, et al. 2013. Structural characterization of H-1 parvovirus: Comparison of infectious virions to empty capsids. J Virol. 87: 5128–5140. ArticlePubMedPMCLink
- Hemminki O, Dos Santos JM, Hemminki A. 2020. Oncolytic viruses for cancer immunotherapy. J Hematol Oncol. 13: 84.ArticlePubMedPMCPDF
- Kashiwakura Y, Tamayose K, Iwabuchi K, Hirai Y, Shimada T, et al. 2005. Hepatocyte growth factor receptor is a coreceptor for adeno-associated virus type 2 infection. J Virol. 79: 609–614. ArticlePubMedPMCLink
- Kaufman HL, Kohlhapp FJ, Zloza A. 2015. Oncolytic viruses: A new class of immunotherapy drugs. Nat Rev Drug Discov. 14: 642–662. ArticlePubMedPMCPDF
- Kontou M, Govindasamy L, Nam HJ, Bryant N, Llamas-Saiz AL, et al. 2005. Structural determinants of tissue tropism and in vivo pathogenicity for the parvovirus minute virus of mice. J Virol. 79: 10931–10943. ArticlePubMedPMCLink
- Lombardo E, Ramirez JC, Garcia J, Almendral JM. 2002. Complementary roles of multiple nuclear targeting signals in the capsid proteins of the parvovirus minute virus of mice during assembly and onset of infection. J Virol. 76: 7049–7059. ArticlePubMedPMCLink
- López-Bueno A, Rubio MP, Bryant N, McKenna R, Agbandje-McKenna M, et al. 2006. Host-selected amino acid changes at the sialic acid binding pocket of the parvovirus capsid modulate cell binding affinity and determine virulence. J Virol. 80: 1563–1573. ArticlePubMedPMCLink
- Mietzsch M, Penzes JJ, Agbandje-McKenna M. 2019. Twenty-five years of structural parvovirology. Viruses. 11: 362.ArticlePubMedPMC
- Nam HJ, Gurda-Whitaker B, Gan WY, Ilaria S, McKenna R, et al. 2006. Identification of the sialic acid structures recognized by minute virus of mice and the role of binding affinity in virulence adaptation. J Biol Chem. 281: 25670–25677. ArticlePubMed
- Paglino J, Tattersall P. 2011. The parvoviral capsid controls an intracellular phase of infection essential for efficient killing of stepwise-transformed human fibroblasts. Virology. 416: 32–41. ArticlePubMedPMC
- Park GS, Best S, Bloom ME. 2005. Two mink parvoviruses use different cellular receptors for entry into CRFK cells. Virology. 340: 1–9. ArticlePubMed
- Parker JS, Murphy WJ, Wang D, O'Brien SJ, Parrish CR. 2001. Canine and feline parvoviruses can use human or feline transferrin receptors to bind, enter, and infect cells. J Virol. 75: 3896–3902. ArticlePubMedPMCLink
- Parker JS, Parrish CR. 1997. Canine parvovirus host range is determined by the specific conformation of an additional region of the capsid. J Virol. 71: 9214–9222. ArticlePubMedPMCLink
- Parrish CR. 2010. Structures and functions of parvovirus capsids and the process of cell infection. Curr Top Microbiol Immunol. 343: 149–176. ArticlePubMed
- Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, et al. 2004. UCSF Chimera—a visualization system for exploratory research and analysis. J Comput Chem. 25: 1605–1612. ArticlePubMed
- Rayet B, Lopez-Guerrero JA, Rommelaere J, Dinsart C. 1998. Induction of programmed cell death by parvovirus H-1 in U937 cells: Connection with the tumor necrosis factor alpha signalling pathway. J Virol. 72: 8893–8903. ArticlePubMedPMC
- Rhode SL 3rd. 1989. Both excision and replication of cloned autonomous parvovirus DNA require the NS1 (rep) protein. J Virol. 63: 4249–4256. ArticlePubMedPMCLink
- Rhode SL 3rd, Paradiso PR. 1983. Parvovirus genome: Nucleotide sequence of H-1 and mapping of its genes by hybrid-arrested translation. J Virol. 45: 173–184. ArticlePubMedPMCLink
- Ruiz Z, Mihaylov IS, Cotmore SF, Tattersall P. 2011. Recruitment of DNA replication and damage response proteins to viral replication centers during infection with NS2 mutants of minute virus of mice (MVM). Virology. 410: 375–384. ArticlePubMed
- Summerford C, Bartlett JS, Samulski RJ. 1999. αVβ5 integrin: A co-receptor for adeno-associated virus type 2 infection. Nat Med. 5: 78–82. ArticlePubMedPDF
- Summerford C, Samulski RJ. 1998. Membrane-associated heparan sulfate proteoglycan is a receptor for adeno-associated virus type 2 virions. J Virol. 72: 1438–1445. ArticlePubMedPMCLink
- Tamura K, Wakimoto H, Agarwal AS, Rabkin SD, Bhere D, et al. 2013. Multimechanistic tumor targeted oncolytic virus overcomes resistance in brain tumors. Mol Ther. 21: 68–77. ArticlePubMed
- Truyen U, Agbandje M, Parrish CR. 1994. Characterization of the feline host range and a specific epitope of feline panleukopenia virus. Virology. 200: 494–503. ArticlePubMed
- Truyen U, Parrish CR. 1992. Canine and feline host ranges of canine parvovirus and feline panleukopenia virus: Distinct host cell tropisms of each virus in vitro and in vivo. J Virol. 66: 5399–5408. ArticlePubMedPMCLink
- Vihinen-Ranta M, Wang D, Weichert WS, Parrish CR. 2002. The VP1 N-terminal sequence of canine parvovirus affects nuclear transport of capsids and efficient cell infection. J Virol. 76: 1884–1891. ArticlePubMedPMCLink
- Vollmers EM, Tattersall P. 2013. Distinct host cell fates for human malignant melanoma targeted by oncolytic rodent parvoviruses. Virology. 446: 37–48. ArticlePubMedPMC
- Ylösmäki E, Cerullo V. 2020. Design and application of oncolytic viruses for cancer immunotherapy. Curr Opin Biotechnol. 65: 25–36. ArticlePubMed
- Zipris D, Hillebrands JL, Welsh RM, Rozing J, Xie JX, et al. 2003. Infections that induce autoimmune diabetes in BBDR rats modulate CD4+CD25+ T cell populations. J Immunol. 170: 3592–3602. ArticlePubMedPDF
References
Supplementary Information
References
Citations
 ePub Link
ePub Link Cite this Article
Cite this Article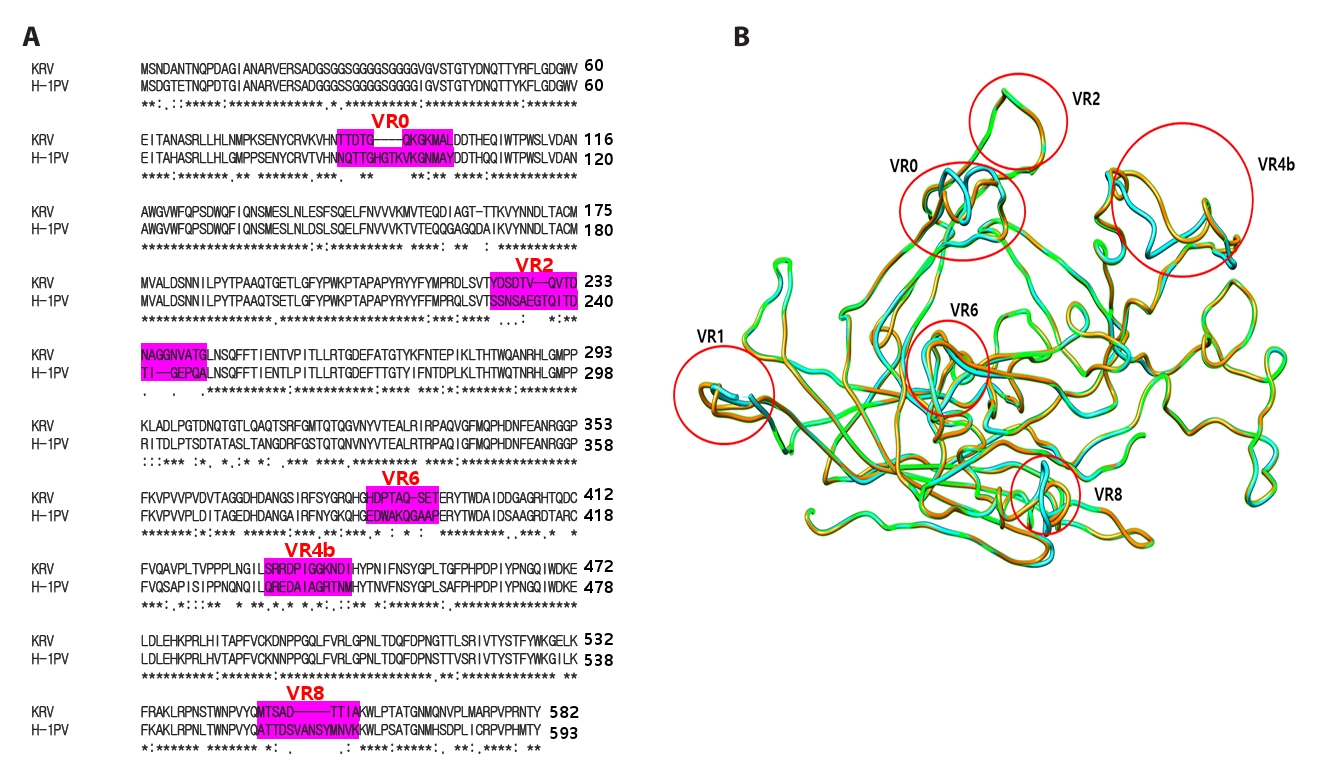
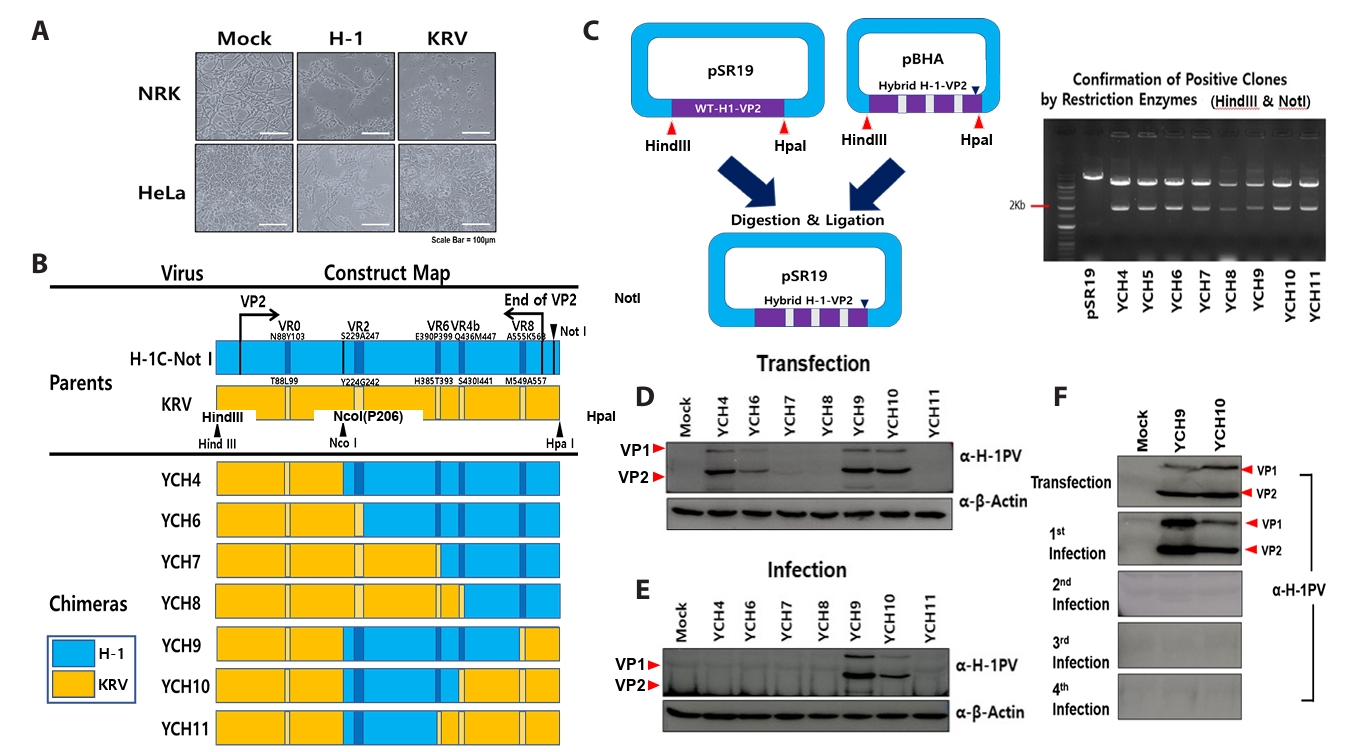
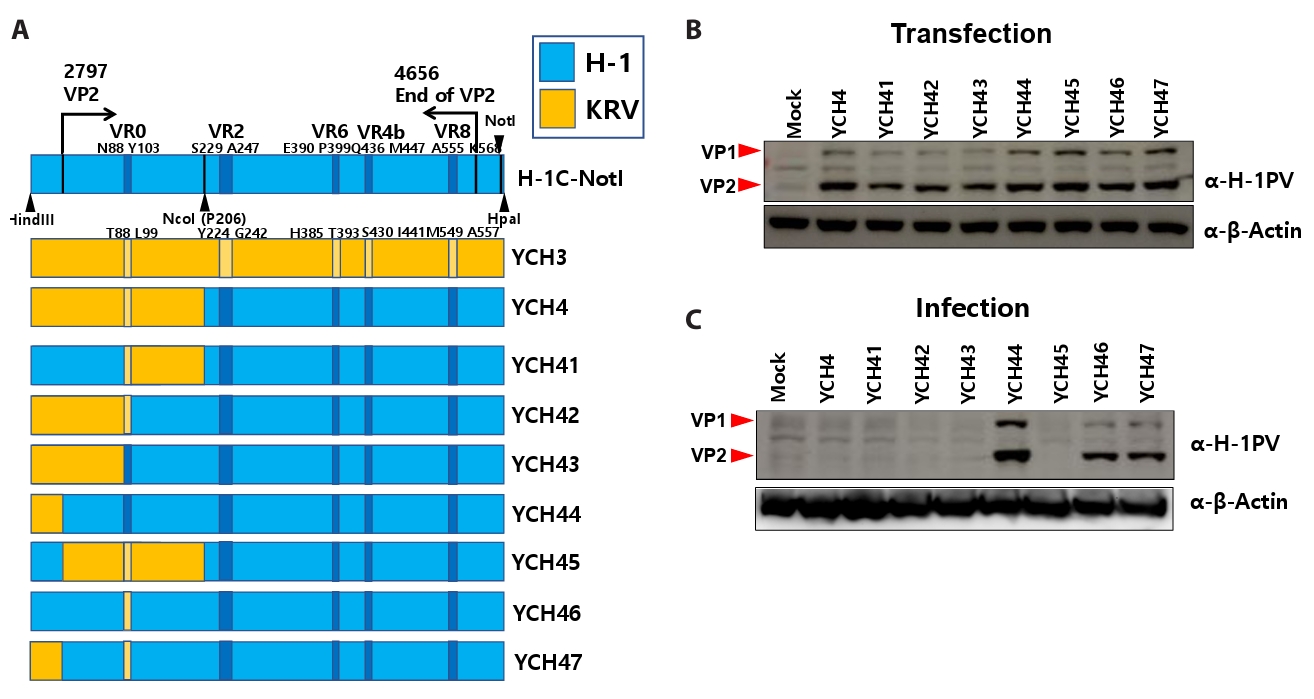
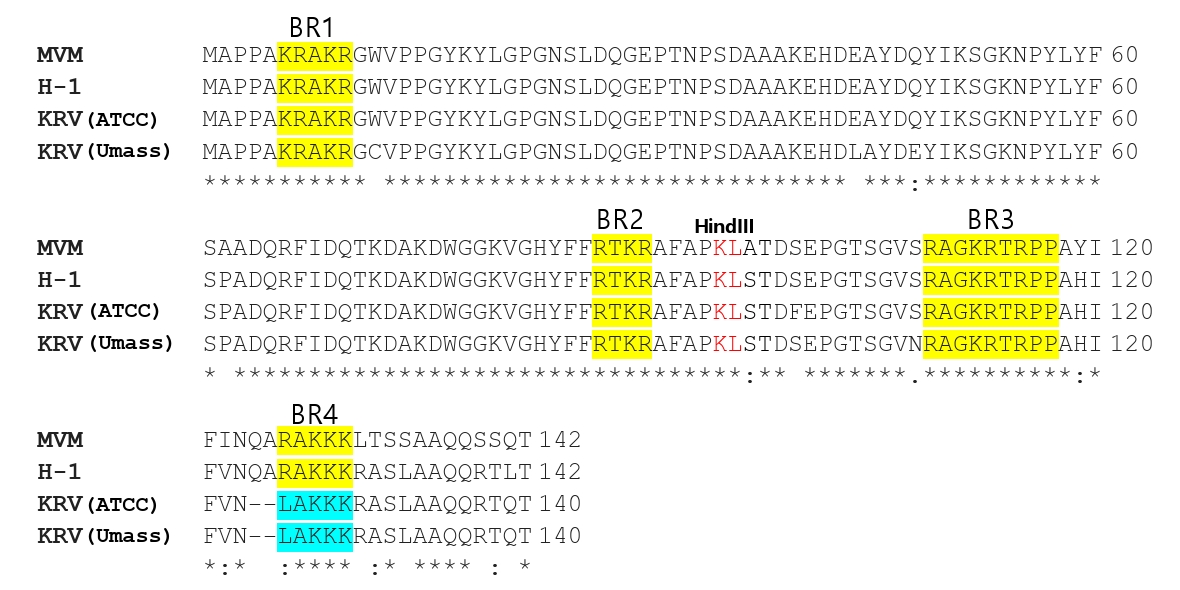
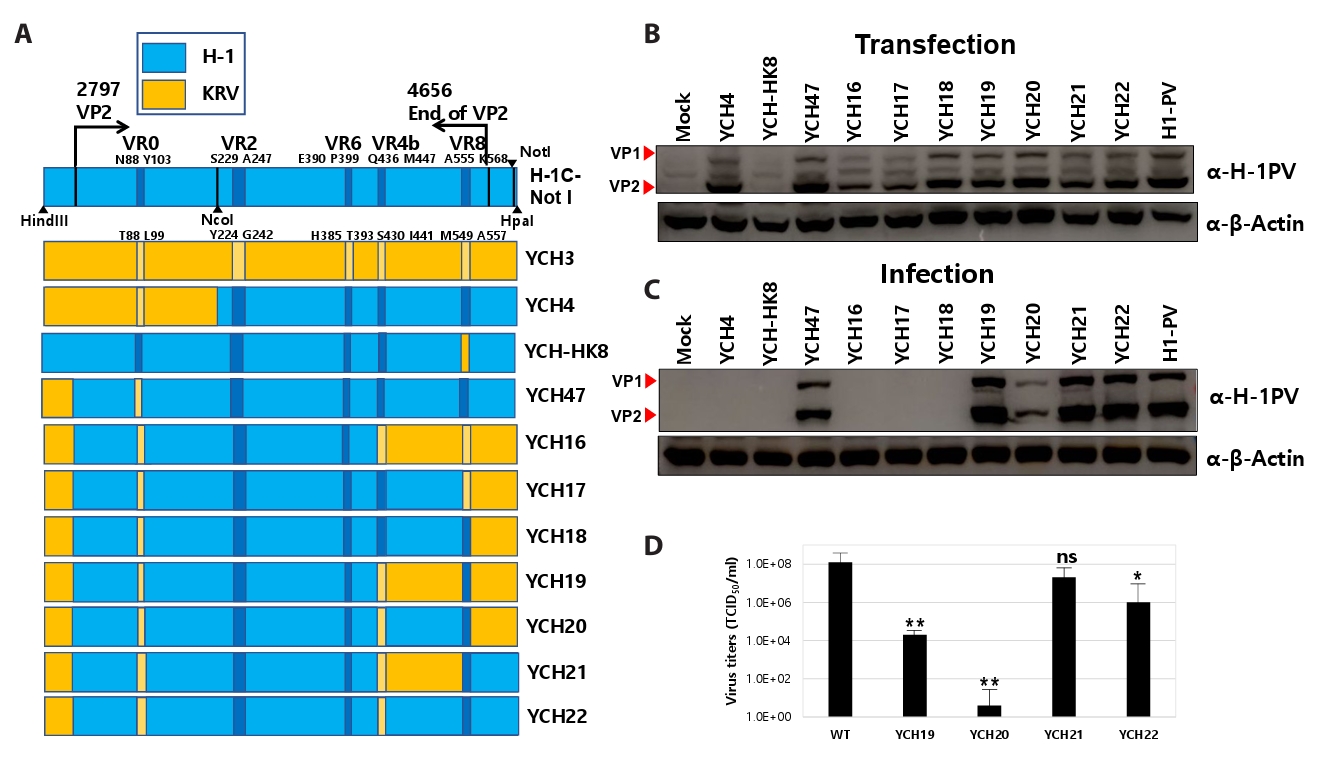
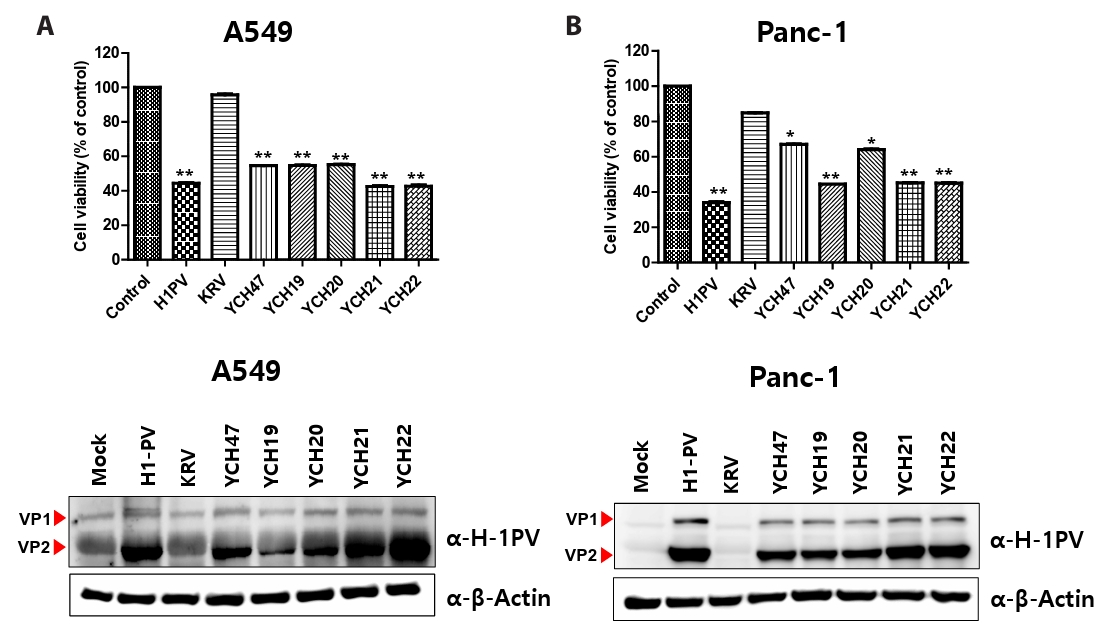






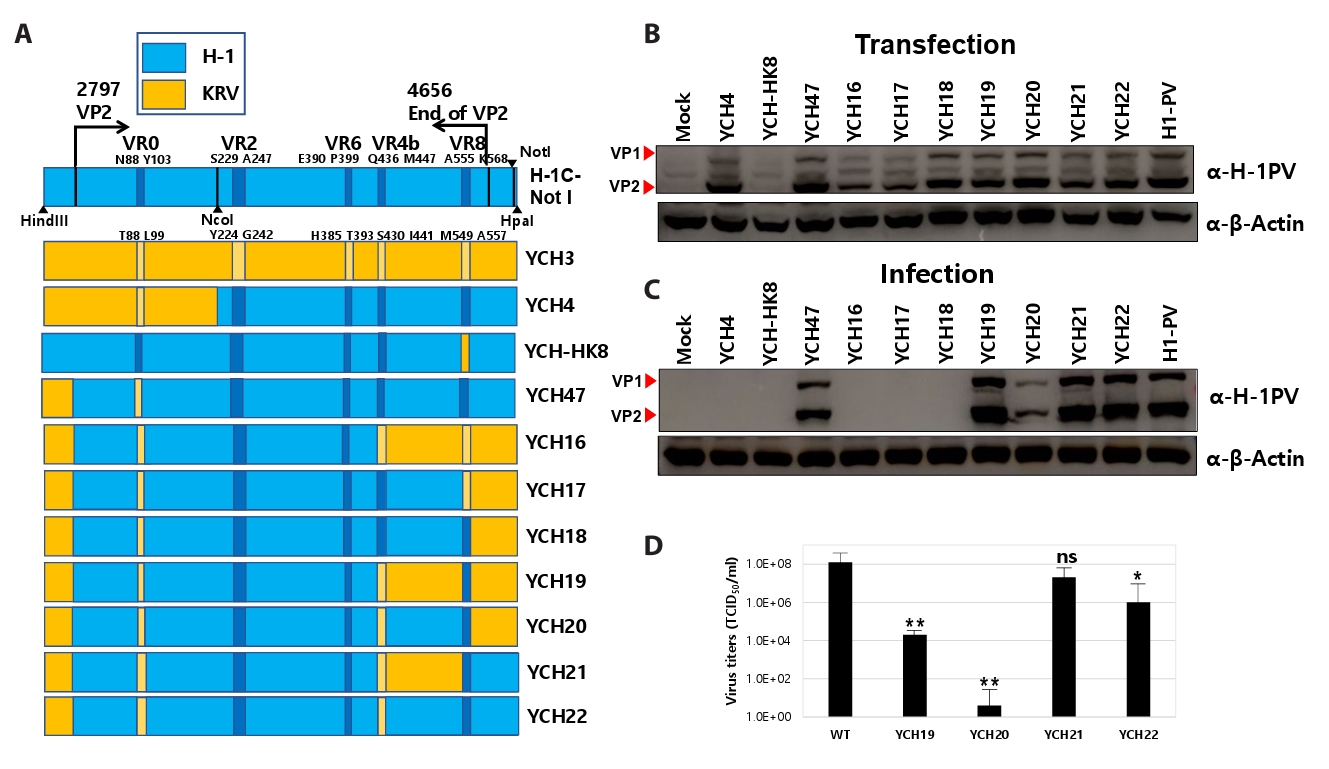
Fig. 1.
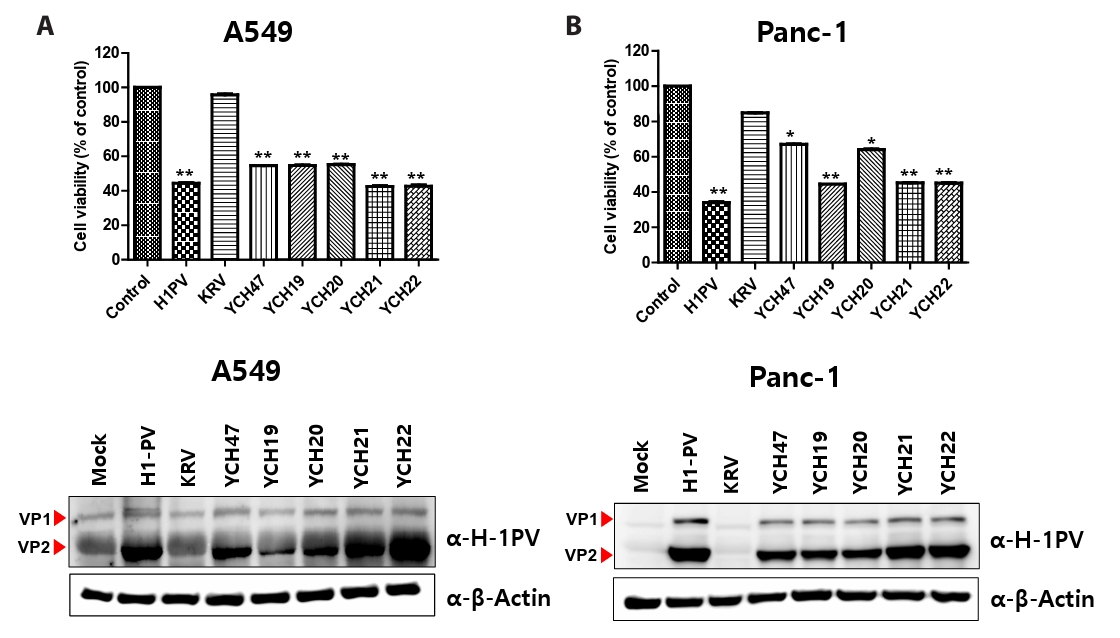
Fig. 2.
Fig. 3.
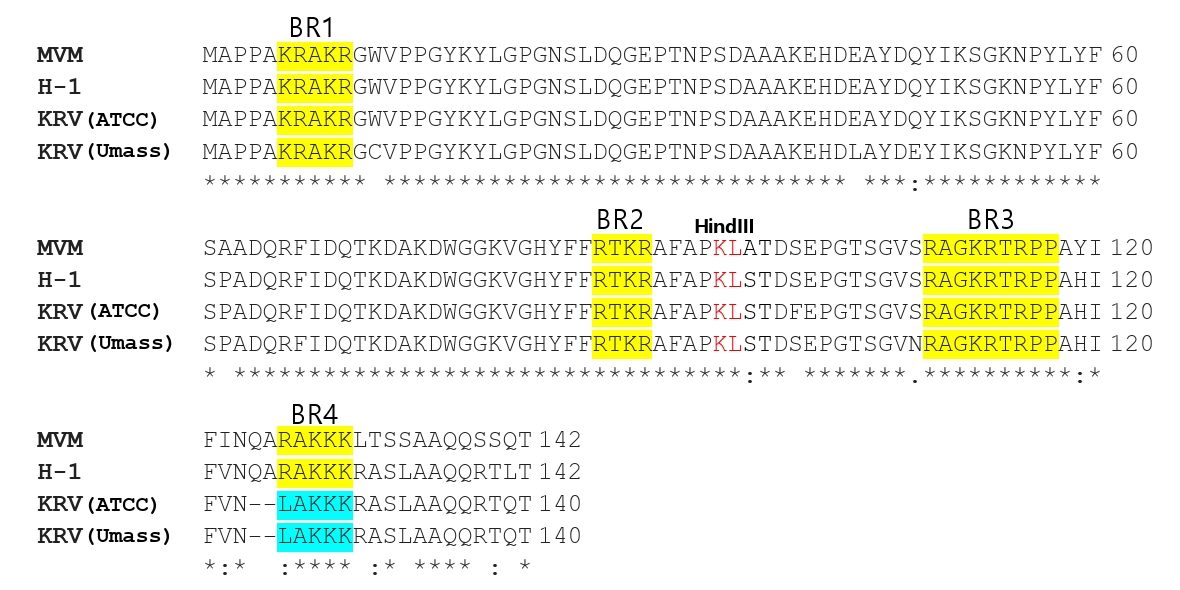
Fig. 4.
Fig. 5.
Fig. 6.
| Genes | Matched nucleotides | Nucleotide homology (%) | Matched amino acids | Amino acid homology (%) |
|---|---|---|---|---|
| NS1 | 2014/2019 | 99.8 | 668/672 | 99.4 |
| NS2 | 564/567 | 99.8 | 186/188 | 98.9 |
| VP1 | 1836/2205 | 86.2 | 594/734 | 78.8 |
| VP2 | 1414/1779 | 83.3 | 443/592 | 73.4 |
Table 1.
TOP

