ABSTRACT
- Marine dinoflagellates are gaining attention as sustainable bioresource for polyunsaturated fatty acids (PUFAs), particularly omega-3 such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). In the present study, we analyzed the FAs and transcriptomic profiles of marine dinoflagellates Amphidinium carterae (D-044) and Prorocentrum minimum (D-127) to evaluate their potential as FAs producers. Gas chromatography-FA methyl ester (GC-FAME) analysis showed that A. carterae is a superior omega-3 producer, yielding a total FA content of 67.6 mg/g DW. DHA accounted for 26.7% of the total FAME profile, which is significantly higher than that of P. minimum (18.1 mg/g DW; DHA 13.1%). Gene Ontology (GO) annotation revealed genes related to FAs and lipid metabolism in A. carterae (1,217 genes) and in P. minimum (2,317 genes), which provide a molecular basis for dinoflagellates with high lipid productivity. Notably, three lipid droplet-associated hydrolase (LDAH) genes with diverse evolutionary origins were identified from A. carterae. These findings suggest a potential expansion of the genetic repertoire related to lipid storage and metabolism, highlighting A. carterae and LDAH as candidates for future biotechnological applications and microalgal metabolic engineering.
-
Keywords: dinoflagellate, Amphidinium, Prorocentrum, fatty acid (FA), omega-3, lipid droplet-associated hydrolase (LDAH)
Introduction
Microalgae are a group of single-cell photosynthetic organisms that occur in diverse aquatic systems from freshwater to the ocean (Singh and Saxena, 2015). Among those, dinoflagellates are one of the largest groups of marine microalgae in terms of species number (Guiry and Guiry, 2026; Taylor et al., 2008). They play a crucial role in primary production and coral reef formation in marine environments (Sampayo et al., 2007). Interestingly, dinoflagellates are known to have complex biosynthetic pathways that yield distinctive metabolites, such as macrolides. For example, several species of Amphidinium and Prorocentrum can synthesize amphidinolides (Kobayashi et al., 2003) and prorocentrolides (Amar et al., 2018). Moreover, some dinoflagellates can produce high levels of fatty acids (FAs), raising commercial interest as aquafeeds and alternatives to fish oil for nutraceuticals (Leblond and Chapman, 2000; Remize et al., 2020).
Fatty acids (FAs) are fundamental components of cellular membranes and storage lipids and serve as precursors to a wide range of metabolites (Ali and Szabó, 2023; De Carvalho and Caramujo, 2018). Polyunsaturated FAs (PUFAs), notably omega-3 and omega-6, play important roles in controlling membrane properties, cell signaling, and inflammation (Calder, 2001). In particular, eicosapentaenoic acid (EPA; C20:5n-3) and docosahexaenoic acid (DHA; C22:6n-3) are representative omega-3, functioning as key regulators of membrane fluidity and cellular defense against stresses (Gorjão et al., 2009; Hu et al., 2024). Also, EPA and DHA are associated with benefits in aging, neurodevelopment, and inflammatory disorder improvements (Hu et al., 2024; Uauy and Dangou, 2006). In this regard, the demand for omega-3 continues to increase, and the global omega-3 market was valued at $2.1–22.8 billion in 2024 (FAO, 2020; Research and Markets, 2021). Their production heavily depends on marine fisheries (Glencross et al., 2025), however, fish oil supply is limited by overfishing and sustainability issues (Byreddy et al., 2019; Tocher et al., 2019). Consequently, microalgae have emerged as sustainable and alternative producers of omega-3 production (Russo et al., 2021).
The dinoflagellates Amphidinium and Prorocentrum are pivotal microalgae in marine ecosystems and are also highly valued as emerging resources for omega-3. Their exceptional capacity to synthesize and store PUFAs positions them as promising and sustainable sources for nutraceutical and pharmaceutical applications (Orefice et al., 2023). However, studies on the omega-3 production potential of dinoflagellates remain fragmented compared to other industrial strains Schizochytrium sp. and Crypthecodinium cohnii. For example, previous studies were often limited to FAs profiling under diverse culture conditions such as temperature (La Motta et al., 2024), light intensity (Molina-Miras et al., 2022; Yin et al., 2023), nutrients (Jiang et al., 2014; Weng et al., 2014), and diverse stresses (Kusumaningtyas et al., 2017; Schmitz et al., 2025). Moreover, the intrinsic omega-3 metabolic flux and storage at the molecular level are poorly understood.
Recent advances in high-throughput sequencing have revealed that the mechanisms involved in FAs biosynthesis in microalgae are complex, with numerous isoforms and unique metabolic flux (Schmitz et al., 2025). Nevertheless, research on dinoflagellates has been limited to transcriptional changes in response to culture conditions and toxins biosynthesis (Bui et al., 2021; Kim et al., 2021, 2023; Muhammad et al., 2026; Wang et al., 2021a). Accordingly, functional genes participating in lipid synthesis in dinoflagellates remain poorly characterized, which is a challenge to solve for future efficient metabolic engineering and commercial process optimization. Therefore, in the present study, we investigated marine dinoflagellates Amphidinium carterae and Prorocentrum minimum, focusing on FAs profiles and transcriptome analysis. Firstly, we observed the lipid droplet formation and quantified FAs using gas chromatography-FA methyl ester (GC-FAME) analysis to compare the profiles. Then, we analyzed transcriptome to identify genes related to FAs metabolisms and lipid droplet formation. The tested species are harmful algal bloom causative dinoflagellates that have been studied for large-scale cultivation and natural compounds production.
Materials and Methods
Cell isolation and strain culture
The strains of A. carterae (D-044) and P. minimum (D-127) used in this study were obtained from the Korea Marine Microalgae Culture Center (Pukyong National University, Korea). Each strain was originally isolated from a single vegetative cell collected from the coastal waters of South Korea, and cultured in f/2 medium without silicate (Ryther and Guillar, 1962). The strains were cultured at 20°C and 65 μmol photons/m2/s of photon flux density under a 12:12 h light–dark cycle. The cells were observed using both light and epifluorescence microscopy (Carl Zeiss Axioskop, Germany) equipped with a ProgRes® CF scan CCD camera (Jenoptik, Germany).
Cell density and size measurements
Changes in cell density was evaluated using a plankton-counting chamber (Matsunami Glass, Japan) under a light microscope (Carl Zeiss Axioskop) to examine the growth patterns of A. carterae and P. minimum. Each sample was fixed by adding Lugol’s solution (final concentration of 1%) and all counts were performed independently in triplicate. Additionally, the average cell size was analyzed using an Auto T4 CellometerTM (Nexcelom Biosciences, USA).
Nile red staining
Nile Red fluorescent dye (CAS No. 7385-67-3; Sigma-Aldrich) was used to stain and observe the neutral lipids in the test species. The stock solution was dissolved as 0.1 mg/ml in acetone (320110; Sigma-Aldrich). Fresh cells (1 × 106 cells/ml) from the exponential, stationary, and decline phases were stained with a final concentration of 1.0 μg/ml. Samples were incubated in darkness for 7 min, and washed three times with fresh f/2 medium. The orange fluorescent was observed using a fluorescence microscope (Carl Zeiss Axioskop) to determine the lipid droplet production.
Fatty acid methyl ester (FAME) analysis
For FA analysis, 5–10 L of the cultures in the exponential phase were harvested by centrifugation at 3,000 × g for 10 min. The supernatant was removed, and the cell pellets were collected and lyophilized through freeze-drying. The freeze-dried biomasses were then precisely weighed to determine the dry cell weight (DCW), which served as the starting materials of FA extraction and analysis. Lipid extractions and transesterification to FAME were performed using 5% acetyl chloride (CH3COCl) in methanol, following the method of Lepage and Roy (1986). FAME profiles of the samples were analyzed using a Shimadzu GC 2010 Plus gas chromatograph (Shimadzu, Japan) with a DB-23 capillary column (Agilent J&W) and a flame ionization detector (FID). Methyl heptadecanoate and a Supelco 37 component FAME Mix (CRM47885, Merck, Germany) were used as the internal standard (IS) and FAME standards, respectively. Detailed analytical conditions were followed previously published methods (Kang et al., 2025). The FAs profile was categorized into saturated FAs (SFAs), monounsaturated FAs (MUFAs), polyunsaturated FAs (PUFAs), and unidentified FAs (UnFAs). The total FAs contents were calculated as mg/g of initial freeze-dried DCW. The FAME content was calculated using the following equation.
RNA extraction and cDNA synthesis
To gather as much gene information as possible, A. carterae and P. minimum cells under diverse conditions were harvested; thermal shock (30°C for 12 and 24 h), exposure to UV irradiation (20 min) and toxic contaminants (0.5 mg/L CuSO4, 0.5 mg/L CuCl2, 0.5 mg/L NaOCl, and 0.5 mg/L NiSO4 for 6 and 24 h), and different growth stages (lag, exponential, and declining phases). About 200 ml of each culture were centrifuged at 3,000 × g for 10 min, and the pellets were mixed with 1 ml of RiboEx solution (Geneall Biotechnology Co. LTD., Korea) and stored at −80°C until the extraction. The total RNA was extracted using GeneAll® Hybrid-RTM miRNA Kit (Geneall) following to the manufacturer’s instructions. RNA quality and quantity were determined using an Agilent 2100 Bioanalyzer (Agilent, USA). The complementary DNA (cDNA) was synthesized using the Illumina® TruSeqTM RNA Sample Preparation Kit (RS122-2001, Illumina Inc., USA). Sequencing was performed using Illumina HiSeq 2500 (Illumina Inc.).
Transcriptome assembly and functional annotation
The raw sequence data was deposited in the National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) database (BioProject accession no. PRJNA14639770). The quality of the raw reads was checked with FastQC_v0.11.7 (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). The reads were cleaned and trimmed by removing adaptor and low-quality reads using Trimmomatic v.0.38. The reads were then assembled using Trinity software and the assemblies were assigned contig codes. Additionally, the CD-HIT program v.4.6 was used for sequence clustering with a default setting to reduce sequence redundancy. All contigs were annotated against public databases gene ontology (GO) with an E-value default cutoff of 1.0E-5. To minimize redundancy of GO annotation terms across the biological process (BP), molecular function (MF), and cellular component (CC) categories, gene assignments were prioritized in the order of BP, MF, and CC. Furthermore, sub-annotations were consolidated into the top categories related to the lipid and fatty acid metabolism.
Identification of LDAH genes and phylogenetic analysis
Identification of lipid droplet-associated hydrolase (LDAH) gene candidates was selected through filtering the annotated transcript to isolate contigs containing conserved domain. The selected contigs were then subjected to a BlastX search to exclude sequences derived from potential contaminations (Bacteria and Fungi). Subsequently, the contigs showing the best hits (E-value < 1.0E-5) within the microalgae or dinoflagellates taxonomy group were retained and designated as the final list of LDAH for subsequent analysis. Amino acid (aa) sequences of LDAH identified from A. carterae and P. minimum were used for the phylogeny analysis. The sequences were aligned using MAFFT. Phylogenetic tree was constructed using the Maximum Likelihood (ML) method with the Whelan and Goldman (WAG) + Freq model with a bootstrap 1,000 replicates in MEGA 6.06 (Tamura et al., 2013).
Results
Morphological, cell growth, and cell size features of A. carterae and P. minimum
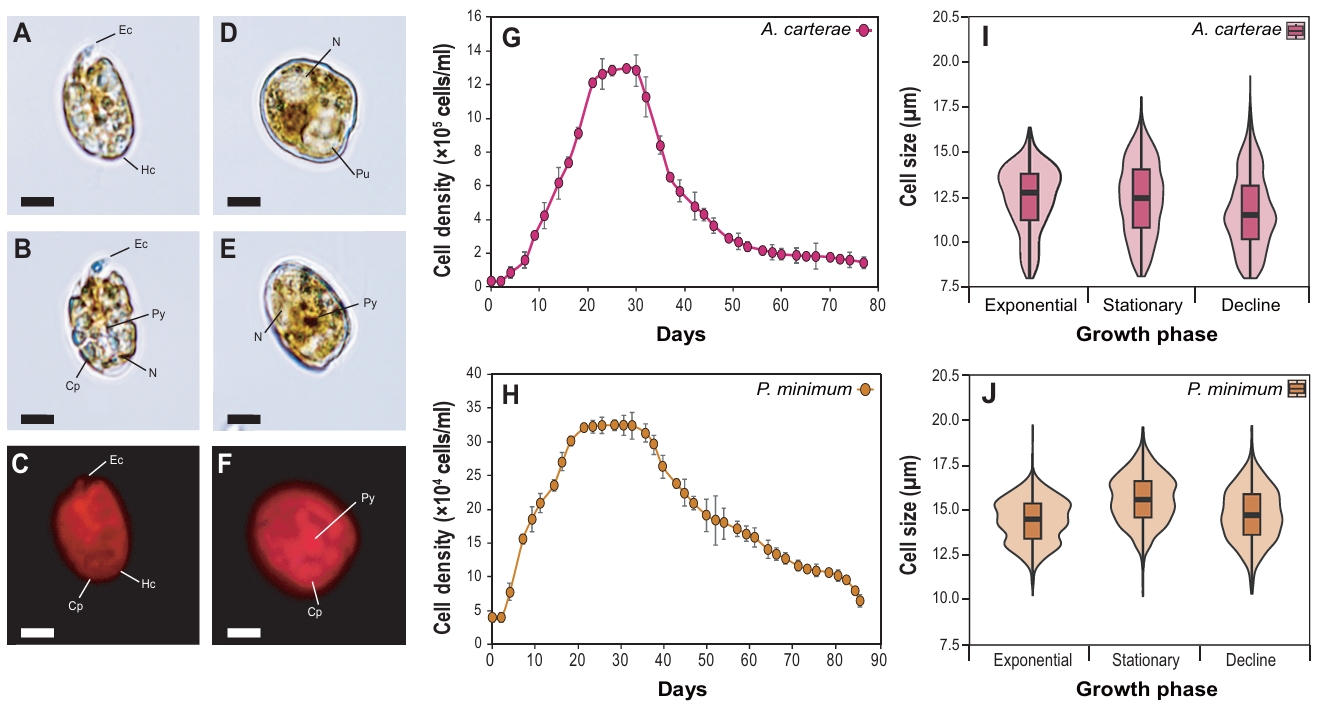
The marine dinoflagellates Amphidinium carterae (D-044) and Prorocentrum minimum (D-127) are unarmored and armored species, respectively (Fig. 1A-1F). The morphology of A. carterae showed an oval shape and dorsoventrally flattened form. Epicone (Ec) was markedly small and tongue-shaped, and Hypocone (Hc) occupied the majority of the cell volume. The nucleus was spherical and primarily positioned in the posterior region of the HC. Chloroplasts (Cps) were spread across the cells, and a ring-shaped pyrenoid was observed. Also, P. minimum (D-127) cells exhibited a typical cordiform shape, enclosed by theca. The nucleus was large and primarily located at the anterior end of the cells. The dark-brown pyrenoid and the plate-shaped Cps were located at the cell center and periphery, respectively.
The strains were cultured under identical conditions at 20°C in f/2 medium to evaluate their growth characteristics, including cell growth patterns (Fig. 1G–1H) and cell size (Fig. 1I–1J). The maximum cell density of A. carterae and P. minimum peaked at 1.29 × 106 cells/ml (day 28) and 3.13 × 105 cells/ml (day 30), respectively. Commonly, the strains started to proliferate on days 7 to 9 after inoculation, and once they reached the stationary phase, a sharp decrease in cell density was observed during the decline phase. Overall cell size of A. carterae was smaller than that of P. minimum. In details, A. carterae exhibited the largest median cell size during the exponential phase (12.6 μm), which decreased to 12.4 μm during the stationary phase and to 11.7 μm during the decline phase. Also, the median cell size of P. minimum was 14.5 μm during the exponential phase and increased to 15.6 μm during the stationary phase. Subsequently, it decreased to 14.7 μm during decline phase.
Total fatty acid methyl esters (FAME) profile
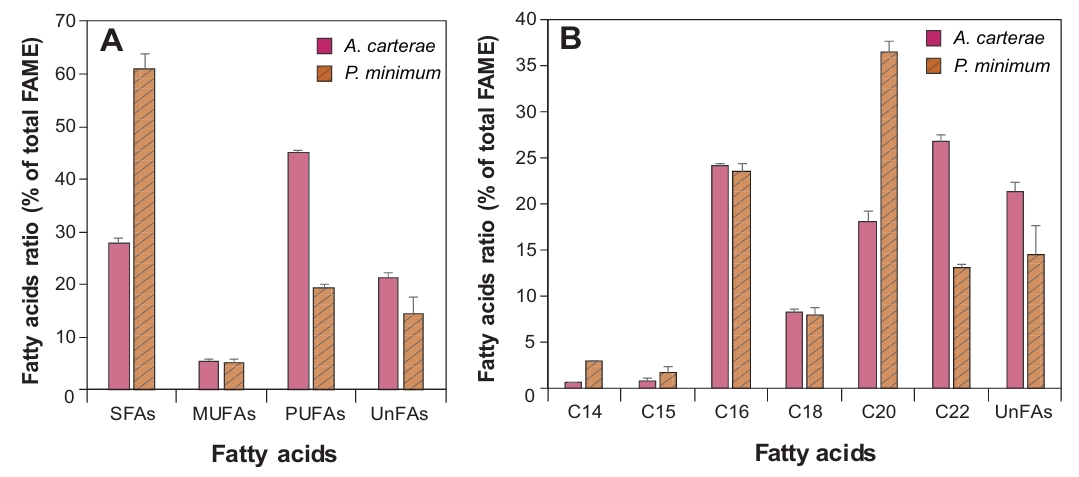
FAs profile of A. carterae and P. minimum was analyzed through FAME analysis (Fig. 2, Table 1). The results showed that SFAs and PUFAs accounted for a high proportion in both species, but there were distinct differences in detailed profiles. Specifically, PUFAs were predominant in A. carterae (45.0%), while P. minimum showed a significantly higher SFA content of 61.0%. In details, palmitic acid (C16:0), which is common to both species, accounted for over 23.5% of total FAs. Among PUFAs, only three omega-3 FAs including alpha-linolenic acid (ALA, C18:3n3), EPA (C20:5n-3), and DHA (C22:6n-3) were detected in both dinoflagellates, and the total concentration in A. carterae (67.6 mg/g DW) was much higher than that of P. minimum (18.1 mg/g DW). The DHA content of A. carterae reached 26.7% of the total FAME profile, approximately double that of P. minimum (13.1%). Conversely, P. minimum contained the highest percentage of SFAs, mainly arachidic acid (C20:0, 32.6%) and palmitic acid (C16:0, 22.5%). In contrast, omega-6 FAs (C18:2n6, C18:3n6, and C20:4n6) were not detected (N.D.) or detected at very low levels (< 1.9%) in both species.
Lipid droplets formation depending on the growth stages
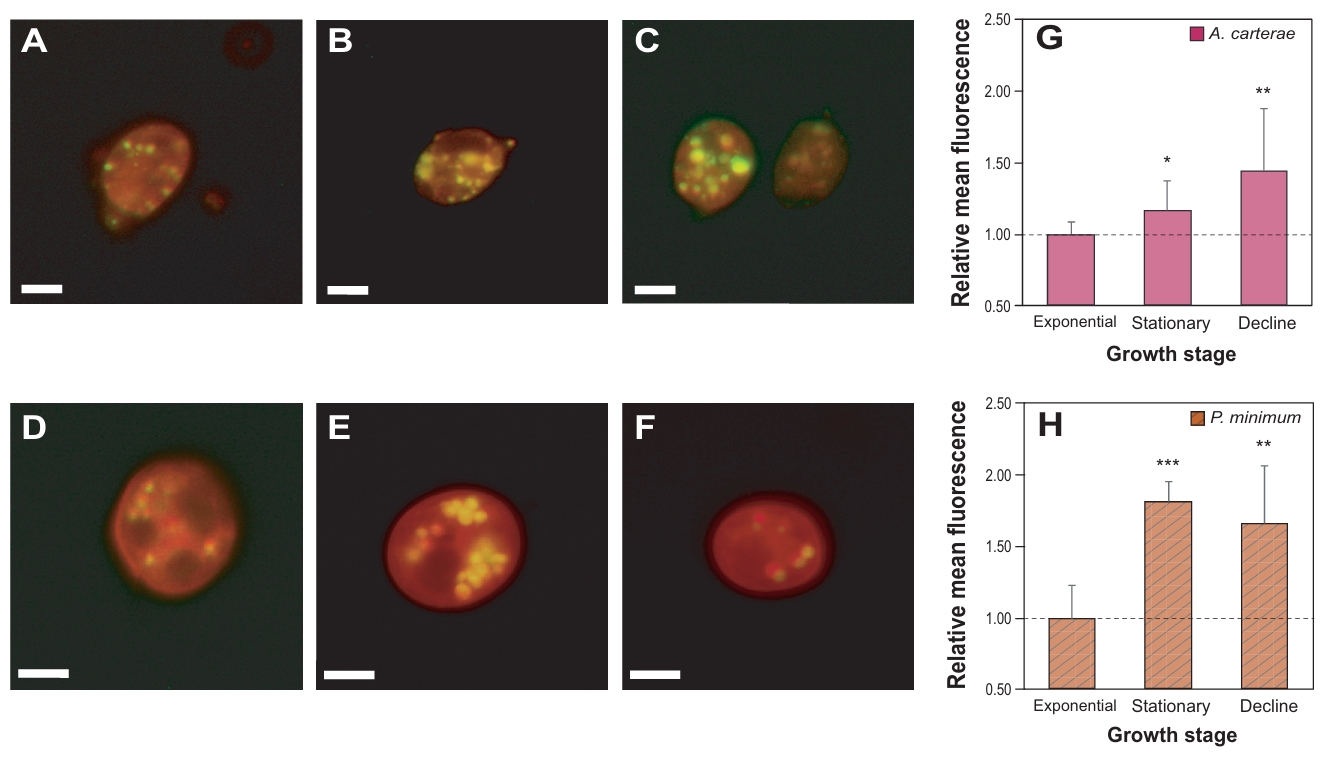
Nile Red staining was performed to observe accumulated neutral lipid in the test species (Fig. 3). The number and size of golden-stained lipid droplets clearly differed depending on the growth phases. In case of A. carterae, the relative mean fluorescence intensity increased along with growth stages, reaching up to 1.4-fold higher in the decline phase compared to the exponential phase (p < 0.01). In contrast, P. minimum peaked a lipid accumulation in the stationary phase (1.8-fold, p < 0.001). The fluorescence intensity decreased slightly at the decline phase, while it remained significantly higher than that of the exponential phase (1.6-fold, p < 0.01).
Transcriptome analysis and lipid related GO annotation
Transcriptome analysis yielded 121,969,762 and 126,793,268 raw reads, corresponding to 12.3 and 12.8 Gb nucleotides in A. carterae and P. minimum, respectively. The total transcripts were clustered into 84,121 and 112,295 transcripts. The transcripts were annotated against the gene ontology (GO) public database (16,066 and 38,233 genes, respectively), and the genes annotated with lipid or FA metabolisms related GO were grouped (Table 2). Overall, P. minimum (2,317 genes) contained a larger number of the transcripts and GO-annotated genes than A. carterae (1,217 genes). However, the proportion of GO terms related to lipid and FA metabolism was higher in A. carterae compared to the total annotated genes. Within the BP category, genes associated with lipid metabolism were 1.8-fold more abundant in P. minimum (1,523 genes, 3.98%) than in A. carterae (818 genes, 5.09%). Similarly, gene categorized in MF were more numerous in P. minimum (779 genes, 1.19%) but showed a higher proportion in A. carterae (384 genes, 2.20%). In the cellular component (CC) category, 15 genes annotated with the ‘lipid droplet’ (GO:0005811) were found in both dinoflagellates.
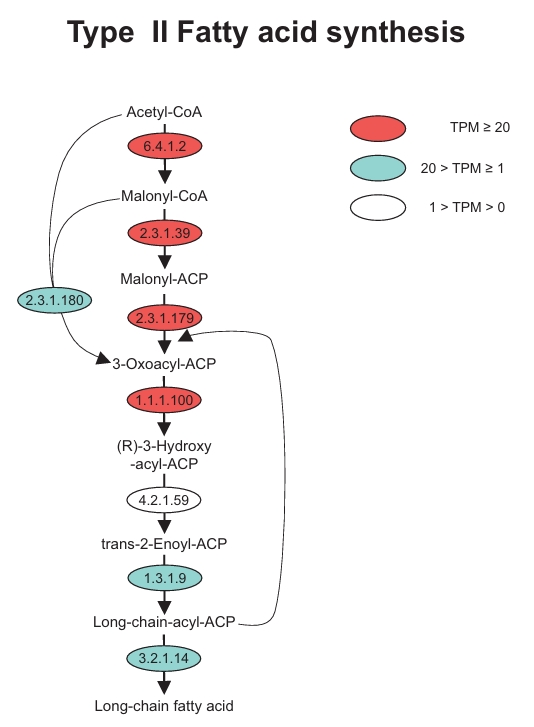
Additionally, we reconstructed the Type II fatty acid synthesis (FAS II) pathway in both dinoflagellates and successfully identified the key enzymes involved in each major stage (Fig. 4). In the initiation stage, acetyl-CoA carboxylase (EC 6.4.1.2) and malonyl-CoA:ACP transacylase (EC 2.3.1.39) exhibited high TPM levels. Within the elongation phase, beta-ketoacyl-ACP synthase (EC 2.3.1.179) and 3-oxoacyl-ACP reductase (EC 1.1.1.100) showed high TPM levels, while beta-ketoacyl-ACP synthase III (EC 2.3.1.180) and enoyl-ACP reductase (EC 1.3.1.9) showed significant TPM levels. For the termination stage, acyl-ACP thioesterase (EC 3.2.1.14), which is responsible for the release of newly synthesized long-chain fatty acids, was also stably expressed.
Phylogenetic analysis of LDAH
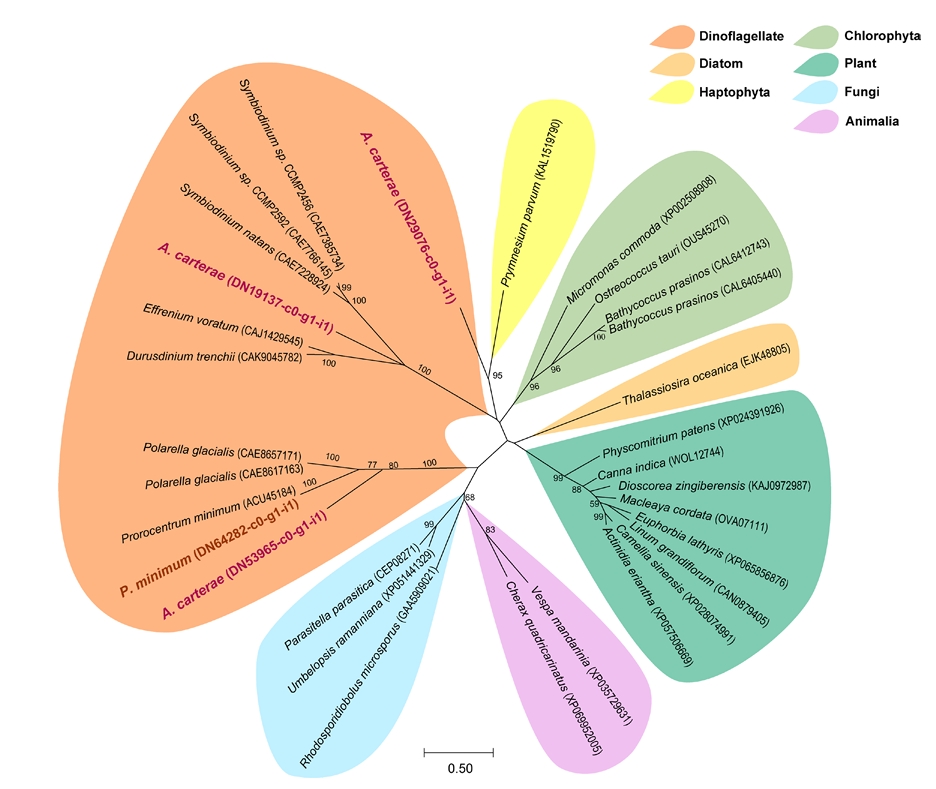
GO analysis revealed a distinct difference in the genes functioning ‘lipid storage’ (GO:0019915) between the species. Four LDAHs were identified including one in P. minimum (DN64282-c0-g1-i1) and three in A. carterae (DN19137-c0-g1-i1, DN29076-c0-g1-i1, and DN53965-c0-g1-i1). Phylogenetic analysis of the selected genes revealed that these belong to the dinoflagellate LDAH lineage or are close to that of haptophyte (Fig. 5). The dinoflagellate LDAHs clade split into two distinct groups: one formed a sister group relationship with the Chlorophyta clade, while the other formed a sister clade with Fungi and Animalia lineages. Meanwhile, the land plant group formed a separate and independent clade, with a high bootstrap support of 99%. Within the dinoflagellate clades, DN19137-c0-g1-i1 from A. carterae clustered closely with the sequences from the dinoflagellates Symbiodinium. Meanwhile, DN29076-c0-g1-i1 from A. carterae was separated from the clade and grouped with the Haptophyta Prymnesium parvum, showing 95% of bootstrap support. Another dinoflagellate clade includes DN53965-c0-g1-i1 from A. carterae and DN64282-c0-g1-i1 from P. minimum, clustering with genes from P. minimum and the dinoflagellate Polarella glacialis, showing 77% of bootstrap support.
Discussion
Marine microalgae account for about half of the total oceanic photosynthetic productivity, playing an important role as a foundation of energy production (Beardall and Raven, 2016). Accordingly, microalgal community transfers energy and biomaterials, especially essential FAs, to higher trophic levels such as fish and invertebrates (Falkowski and Raven, 2007). Also, change in FAs composition contribute to regulating cell membrane fluidity and osmotic pressure and maintaining pigments stability, thereby protecting the cells even in extreme environments (Brett and Müller-Navarra, 1997). Several studies have reported that dinoflagellates possess distinct FAs composition, synthesis pathway, and storage form (Remize et al., 2020). Especially, dinoflagellates store higher levels of FAs with enriched PUFAs compared to other microalgae. Of them, the genera Amphidinium and Prorocentrum are known to accumulate high concentrations of omega-3 (Jaeckisch et al., 2011; Řezanka et al., 2017).
In the present study, the total FAs yield of A. carterae was 67.6 mg/g DW, more than 3.7 times higher than that of P. minimum (18.1 mg/g DW), showing a clear advantage in the productivity. FAs yield of A. carterae was similar to that of industrial strains Nannochloropsis oculata (50–60 mg/g DW) (Ma et al., 2016) and Isochrysis galbana (40–45 mg/g DW) (Ryckebosch et al., 2012). Regarding PUFAs, consistent with our results, the PUFA content of A. carterae was reported to be as high as 53.2%, with DHA and EPA accounting for 30.6% and 19.7%, respectively (Mendoza-Flores and Sánchez-Saavedra, 2023). These values are significantly higher than those reported for other dinoflagellates. For example, DHA levels of only 8–23% have been observed in marine gymnodinioid dinoflagellates (Karenia, Karlodinium, and Takayama) (Mooney et al., 2007). Also, Prorocentrum micans was reported to contain 22.8% of DHA and 13.1% of EPA, both of which were higher than those of P. minimum in the present study (Suh et al., 2015). Furthermore, while PUFAs accounted for up to 60% of total FA in marine green algae Tetraselmis suecica and diatom Phaeodactylum tricornutum, but their DHA remained minimal ranging from 0.2 to 1.7% (Qiao et al., 2016; Volkman et al., 1989). These findings demonstrate that although total FA yield and PUFA profiles vary by species and strains (Vazhappilly and Chen, 1998), dinoflagellates nevertheless showed a remarkable capacity for PUFA production.
Recent research on dinoflagellates has focused on transcriptome analysis due to their enormous and condensed genomic structure (Bui et al., 2025; Muhammad et al., 2025; Wang et al., 2021b). Number of functional genes were identified and their molecular responses were elucidated under various environmental stressors (Abassi and Ki, 2022; Abassi et al., 2023), while only a few studies have analyzed FAs related genes. In this regard, we identified genes participating in the lipid and FAs metabolisms through transcriptome analysis in the two marine dinoflagellates. Through GO analysis, the number of transcripts, 1,217 in A. carterae and 2,317 in P. minimum, were identified. In other cases, a study on Heterocapsa bohaiensis revealed significant changes in transcripts after treatment with allelochemicals, and the key gene groups involved in PUFA production (Acetyl-CoA carboxylase, FA desaturase, and elongase) were significantly up-regulated (Bachvaroff et al., 2014). Also, Kalinina et al. (2025) reported that expression of FA desaturase was suppressed under phosphorus-limited conditions, accompanied by a decrease in UFAs production in Prorocentrum cordatum. These results suggest that the FA metabolism responds sensitively to environmental changes (Chang et al., 2026; Thiyagarajan et al., 2020). In this regard, comparative transcriptomic analysis among dinoflagellates revealed species-specific differences in the number of FAs-associated genes and their gain/loss patterns (Stephens et al., 2021). Consistently, Rein et al. (2025) reported that while K. brevis possesses a high number of isoforms, proportion of core FA gene families was relatively low compared to others. This aligns with our findings, where the proportion of FAs related genes was markedly lower in P. minimum. These suggest that P. minimum contains a larger transcript due to various isoforms and gene replication, whereas A. carterae exhibits a more enriched genetic profile in terms of FA metabolisms.
When analyzing GO terms related to FAs metabolism, we focused on lipid storage (GO:0019915) category. Since PUFAs are stored in LD in the form of triglycerides (TAG) as well as cell membranes, genes participating in LD formation and maintenance also play an important role as lipid regulators (Danielli et al., 2023). In this regard, LDAH plays a crucial role in maintaining lipid homeostasis by regulating the balance between FA degradation and storage within LD (Wilfling et al., 2014). Although LDAH possesses a hydrolase domain, current research has suggested that it promotes LD fusion and induces intracellular TAG accumulation rather than hydrolysis (Goo et al., 2014, 2017; Kong et al., 2018; Kory et al., 2017). In details, biochemical analyses showed that LDAH displayed weak cholesterol ester hydrolase activity but little TAG hydrolase activity (Goo et al., 2014, 2017). The enzyme and coding genes were primarily identified in model land plants (Zhao et al., 2025), where the presence of LDAH-like proteins was first identified in the LD of the marine protist Aurantiochytrium limacinum (Yoneda et al., 2022). Interestingly, we identified three distinct LDAHs in A. carterae, whereas only a single gene was identified in P. minimum. Notably, phylogenetic analysis revealed that the LDAHs had a distinct evolutionary origin, implying an evolutionary strategy that enables them to gain functional flexibility for environmental adaptation. Indeed, Goold et al. (2015) also reported that various microalgal species possess evolutionarily unique LD-associated protein families, which perform complementary roles in lipid redistribution, storage, and release. The present study analyzed FA profiles and associated transcripts to identify LDAHs in dinoflagellates for the first time. Furthermore, the multigenic LDAH system of A. carterae can be utilized in metabolic engineering research to improve lipid productivity and/or storage in microalgae with industrial potential. In this regard, functional studies are needed to experimentally verify the enzymatic role and specific regulatory mechanisms of LDAH system in marine dinoflagellates.
Conclusion
The present study provides a molecular and biochemical characterization of FA metabolism in two marine dinoflagellates A. carterae and P. minimum. Through Nile red-based LD observation and GC-FAME analyses, we demonstrated that A. carterae possesses a superior capacity for omega-3 (DHA and EPA) biosynthesis and lipid storage. Also, transcriptomic analysis revealed an enriched genes associated with lipid and FAs metabolism in A. carterae, providing a molecular basis for its robust biosynthetic potential. The three LDAHs identified in A. carterae suggest significant gene diversification that is likely to contribute to lipid storage and related metabolic flexibility. Collectively, these findings imply that A. carterae harbors an enhanced and flexible genetic system for lipid production and highlight that A. carterae and LDHA are noteworthy candidates for blue biotechnology applications and genetic engineering research.
Acknowledgments
We would like to thank Mr. Taehee Kim for microscopic observations and Dr. Buhari Lawan Muhammad for critical comments on an earlier version of this paper.
Author Contributions
Han-Sol Kim and Jang-Seu Ki contributed to the study conception and design. Material preparation, data collection and analysis were performed by Han-Sol Kim, Su-Hwa Lee, Zhun Li, and Hah Young Yoo. The first draft of the manuscript was written by Han-Sol Kim and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Conflict of Interest
The authors declare no conflict of interests.
Funding
This research was funded by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (RS-2024-00354842 and RS-2024-00359501).
Data Availability
The datasets presented in this study can be found in online NCBI Sequence Read Archive (SRA) database (PRJNA14639770).
Ethical Statements
Not applicable.
Fig. 1.Light and fluorescence microscopy images of the marine dinoflagellates Amphidinium carterae (A–C) and Prorocentrum minimum (D–F). Ec, epicone; Hc, hypocone; Cp, chloroplast; Py, pyrenoid; Pu, pusule; N, nucleus. Scale bars represent 10 µm. Growth curve and cell size of the marine dinoflagellates A. carterae (G–H) and P. minimum (I–J).
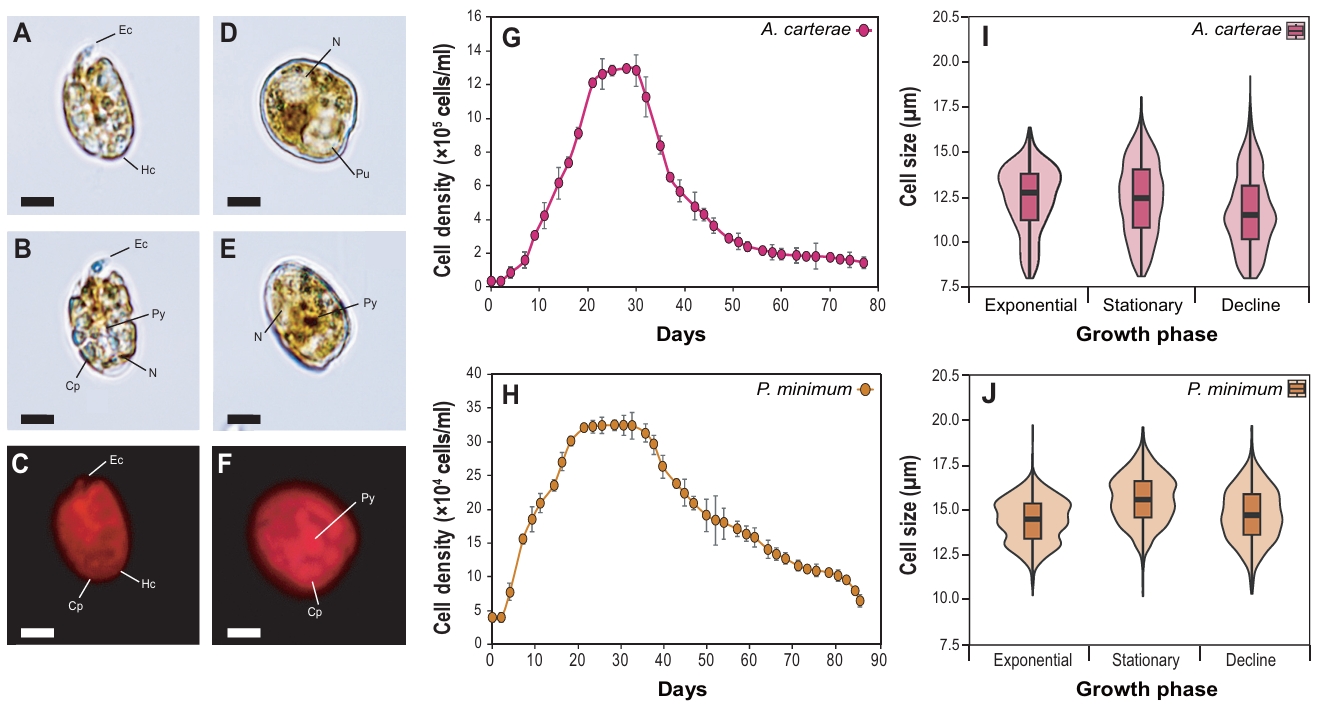
Fig. 2.Comparison of fatty acid methyl esters (FAME) compositions (% of total FAME) in the marine dinoflagellates Amphidinium carterae and Prorocentrum minimum. Percentages of saturated FAs (SFAs), monounsaturated FAs (MUFAs), polyunsaturated FAs (PUFAs), and unidentified FAs (UnFAs) in the total FAs (A). Compositional FMAE profile of total FAs (B).
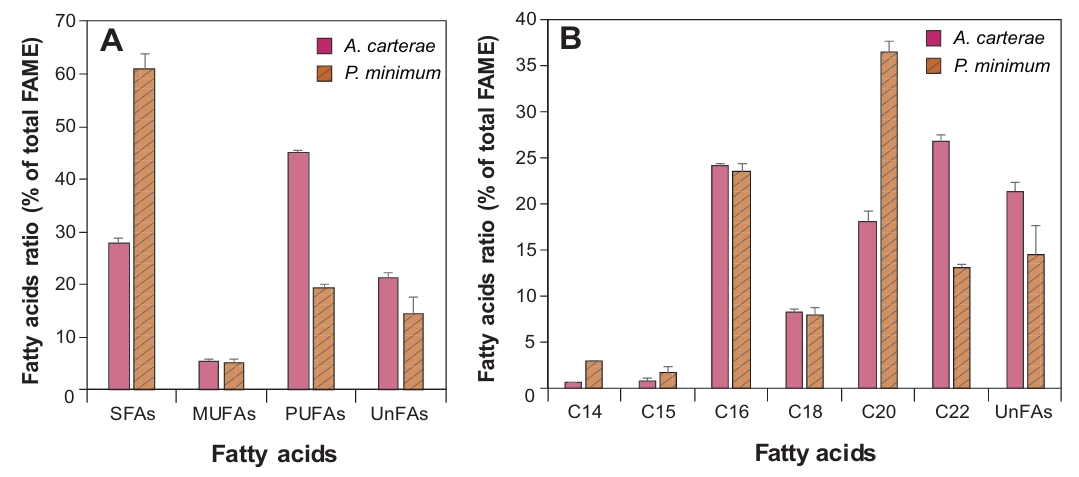
Fig. 3.Nile red staining of the marine dinoflagellates Amphidinium carterae (A–C) and Prorocentrum minimum (D–F). Algal cells with lipid droplets as shown in golden color, and each figure was taken under different growth stages; exponential (A, D), stationary (B, E), and decline phase (C, F). Scale bars represent 10 µm. Relative mean fluorescent levels of lipid droplets in A. carterae (G) and P. minimum (H) under the different growth stages.
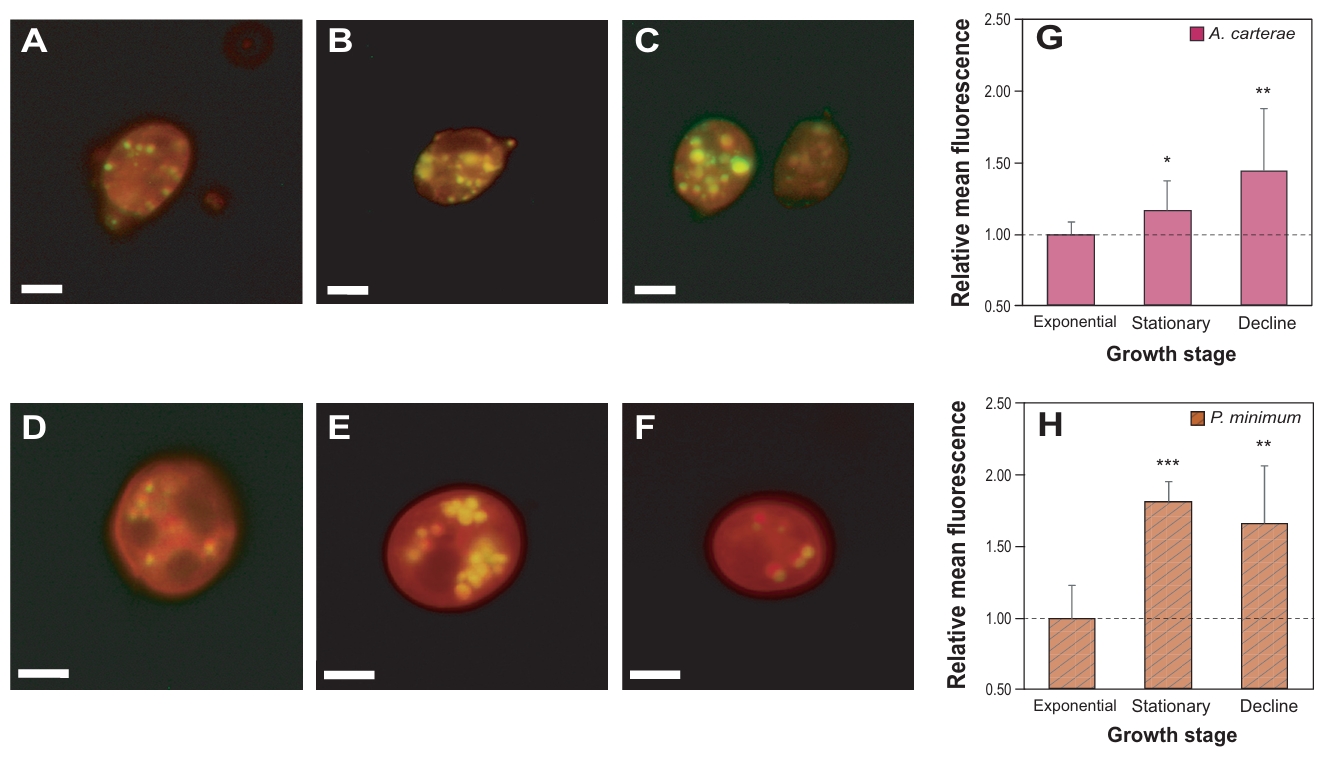
Fig. 4.Identification of genes involved in the Type II fatty acid synthesis (FAS II) in marine dinoflagellates Amphidinium carterae and Prorocentrum minimum. The level of Transcripts Per Million (TPM) is indicated in red, green, and white.
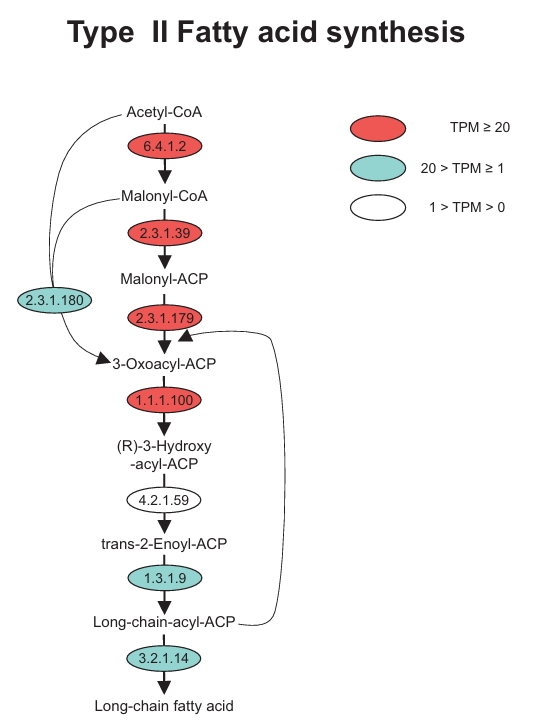
Fig. 5.Phylogenetic tree of lipid droplet-associated hydrolase (LDAH) proteins identified in the marine dinoflagellates Amphidinium carterae and Prorocentrum minimum.
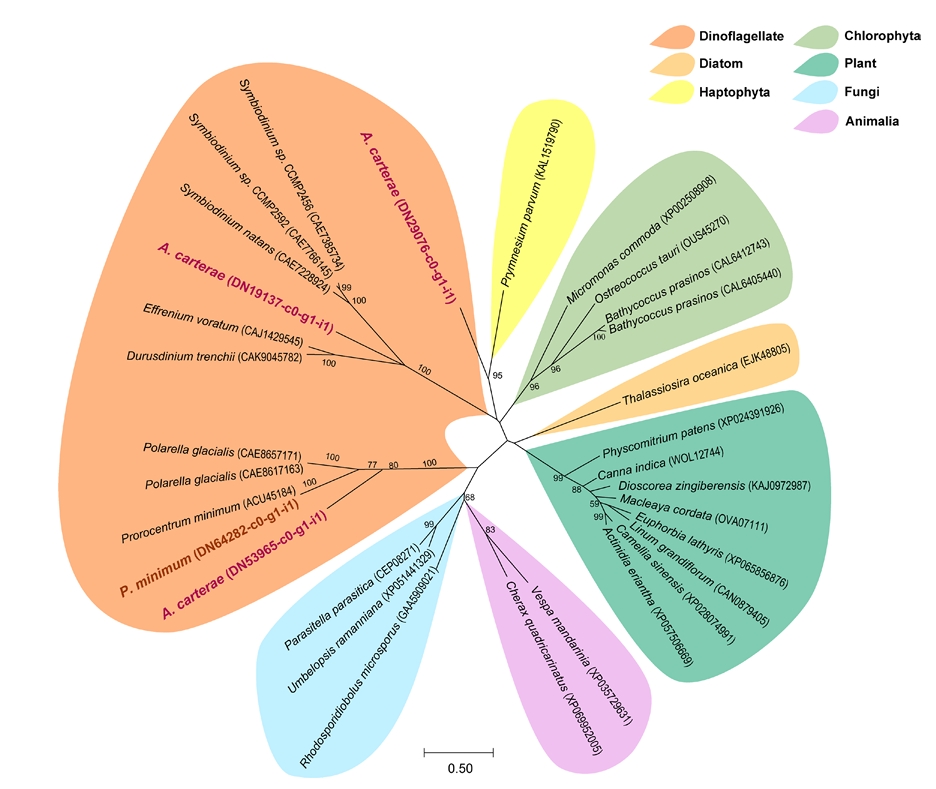
Table 1.Comparison of fatty acid yields (mg/g DW and % DW) and FAME profiles (%) in marine dinoflagellates Amphidinium carterae and Prorocentrum minimum. DW, dry weight; FAME, fatty acid methyl esters; N.D., not detected. Values are expressed as Mean ± SD (n = 3).
|
Contents |
FAs yield (mg/g DW) |
FAs yield (DW %) |
FAME profile (%) |
|
A. carterae
|
P. minimum
|
A. carterae
|
P. minimum
|
A. carterae
|
P. minimum
|
|
C14:0 |
1.0 ± 0.1 |
3.0 ± 0.5 |
0.1 ± 0.0 |
0.3 ± 0.1 |
0.6 ± 0.0 |
2.9 ± 0.1 |
|
C14:1 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C15:1 |
1.1 ± 0.5 |
1.6 ± 0.7 |
0.1 ± 0.0 |
0.2 ± 0.1 |
0.8 ± 0.3 |
1.7 ± 0.8 |
|
C16:0 |
35.8 ± 3.6 |
23.4 ± 3.6 |
3.6 ± 0.4 |
2.3 ± 0.4 |
23.3 ± 0.4 |
22.5 ± 0.8 |
|
C16:1 |
1.3 ± 0.2 |
1.1 ± 0.2 |
0.1 ± 0.0 |
0.1 ± 0.0 |
0.8 ± 0.1 |
1 ± 0.0 |
|
C18:0 |
4.1 ± 0.3 |
3.1 ± 0.5 |
0.4 ± 0.0 |
0.3 ± 0.1 |
2.7 ± 0.2 |
3.1 ± 0.8 |
|
C18:1 cis n9 |
6.1 ± 1.6 |
2.1 ± 0.5 |
0.6 ± 0.2 |
0.2 ± 0.0 |
3.9 ± 0.5 |
2 ± 0.1 |
|
C18:1 trans n9 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C18:2 cis n6 |
N.D. |
0.1 ± 0 |
N.D. |
N.D. |
N.D. |
0.1 ± 0.0 |
|
C18:2 trans n6 |
2.0 ± 0.3 |
1.8 ± 0.4 |
0.2 ± 0.0 |
0.2 ± 0.0 |
1.3 ± 0.1 |
1.7 ± 0.1 |
|
C18:3n6 |
N.D. |
0.1 ± 0.0 |
N.D. |
N.D. |
N.D. |
0.1 ± 0.0 |
|
C18:3n3 |
0.6 ± 0.1 |
0.9 ± 0.1 |
0.1 ± 0.0 |
0.1 ± 0.0 |
0.4 ± 0.1 |
0.9 ± 0.1 |
|
C20:0 |
2.0 ± 0.2 |
33.9 ± 5.1 |
0.2 ± 0.0 |
3.4 ± 0.5 |
1.3 ± 0.2 |
32.6 ± 1.3 |
|
C20:1n9 |
N.D. |
0.5 ± 0.1 |
N.D. |
N.D. |
N.D. |
0.5 ± 0.0 |
|
C20:2 |
N.D. |
0.1 ± 0.1 |
N.D. |
N.D. |
N.D. |
0.1 ± 0.0 |
|
C21:0 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C20:3n6 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C20:4n6 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C20:3n3 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C20:5n3 |
25.7 ± 1.2 |
3.5 ± 1.0 |
2.6 ± 0.1 |
0.3 ± 0.1 |
16.8 ± 1.0 |
3.3 ± 0.4 |
|
C22:0 |
0.2 ± 0.2 |
N.D. |
N.D. |
N.D. |
0.1 ± 0.2 |
N.D. |
|
C22:1n9 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C23:0 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
C22:6n3 |
41.3 ± 6.1 |
13.7 ± 2.9 |
4.1 ± 0.6 |
1.4 ± 0.3 |
26.7 ± 0.8 |
13.1 ± 0.4 |
|
C24:1 |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
N.D. |
|
Unidentified FAs |
33.1 ± 5.1 |
15.7 ± 6.3 |
3.3 ± 0.5 |
1.6 ± 0.6 |
21.4 ± 1.0 |
14.5 ± 3.2 |
Table 2.Gene Ontology (GO) classification of lipid and fatty acid metabolism-related genes identified from marine dinoflagellates Amphidinium carterae and Prorocentrum minimum. The percentages of each GO category represent the ratio of the number of genes selected within the category. N.D., not detected.
|
GO category |
Top GO |
GO term |
No. genes in A. carterae
|
No. genes in P. minimum
|
|
Biological process |
GO:0016042 |
lipid catabolic process |
170 |
220 |
|
GO:0006631 |
fatty acid metabolic process |
191 |
240 |
|
GO:0046486 |
glycerolipid metabolic process |
155 |
285 |
|
GO:1901568 |
fatty acid derivative metabolic process |
10 |
36 |
|
GO:0008610 |
lipid biosynthetic process |
48 |
139 |
|
GO:0006644 |
phospholipid metabolic process |
17 |
62 |
|
GO:0006643 |
membrane lipid metabolic process |
104 |
214 |
|
GO:0006869 |
lipid transport |
43 |
168 |
|
GO:1903509 |
liposaccharide metabolic process |
40 |
34 |
|
GO:0008202 |
steroid metabolic process |
18 |
91 |
|
GO:0030258 |
lipid modification |
3 |
19 |
|
GO:0055091 |
phospholipid homeostasis |
N.D. |
3 |
|
GO:0006720 |
isoprenoid metabolic process |
11 |
10 |
|
GO:0019216 |
regulation of lipid metabolic process |
2 |
1 |
|
GO:0019915 |
lipid storage |
6 |
1 |
|
|
5.09% |
3.98% |
|
Cellular component |
GO:0005811 |
lipid droplet |
15 |
15 |
|
|
0.10 % |
0.05 % |
|
Molecular function |
GO:0016407 |
acetyltransferase activity |
67 |
85 |
|
GO:0016705 |
oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen |
205 |
587 |
|
GO:0120515 |
fatty acid-CoA ligase activity |
33 |
39 |
|
GO:0004312 |
fatty acid synthase activity |
71 |
59 |
|
GO:0009922 |
fatty acid elongase activity |
8 |
9 |
|
|
2.20% |
1.19% |
References
- Abassi S, Ki JS. 2022. Increased nitrate concentration differentially affects cell growth and expression of nitrate transporter and other nitrogen-related genes in the harmful dinoflagellate Prorocentrum minimum. Chemosphere. 288: 132526.ArticlePubMed
- Abassi S, Kim HS, Bui QTN, Ki JS. 2023. Effects of nitrate on the saxitoxins biosynthesis revealed by sxt genes in the toxic dinoflagellate Alexandrium pacificum (group IV). Harmful Algae. 127: 102473.ArticlePubMed
- Ali O, Szabó A. 2023. Review of eukaryote cellular membrane lipid composition, with special attention to the fatty acids. Int J Mol Sci. 24: 15693.ArticlePubMedPMC
- Amar M, Aráoz R, Iorga BI, Yasumoto T, Servent D, et al. 2018. Prorocentrolide-A from cultured Prorocentrum lima dinoflagellates collected in Japan blocks sub-types of nicotinic acetylcholine receptors. Toxins. 10: 97.ArticlePubMedPMC
- Bachvaroff TR, Handy SM, Place AR, Delwiche CF. 2014. Comparative transcriptomics reveals common gene content but high sequence divergence in dinoflagellates. PLoS One. 9: e94012.ArticlePubMedPMC
- Beardall J, Raven JA. 2016. Carbon acquisition by microalgae. In Borowitzka MA, Beardall J, Raven JA. (eds.), The Physiology of Microalgae, pp. 89–99. SpringerArticle
- Brett MT, Müller-Navarra DC. 1997. The role of highly unsaturated fatty acids in aquatic foodweb processes. Freshw Biol. 38: 483–499. ArticleLink
- Bui QTN, Kim T, Kim HS, Ki JS. 2025. Defensive responses of most antioxidant genes in the freshwater dinoflagellate Palatinus apiculatus to cadmium stress and their implications. Ecotoxicol Environ Saf. 289: 117380.ArticlePubMed
- Bui QTN, Kim H, Park H, Ki JS. 2021. Salinity affects saxitoxins (STXs) toxicity in the dinoflagellate Alexandrium pacificum, with low transcription of SXT-biosynthesis genes sxtA4 and sxtG. Toxins. 13: 733.ArticlePubMedPMC
- Byreddy AR, Yoganantharjah P, Gupta A, Gibert Y, Puri M. 2019. Suitability of novel algal biomass as fish feed: Accumulation and distribution of omega-3 long-chain polyunsaturated fatty acid in zebrafish. Appl Biochem Biotechnol. 188: 112–123. ArticlePubMedPDF
- Calder PC. 2001. Polyunsaturated fatty acids, inflammation, and immunity. Lipids. 36: 1007–1024. ArticlePubMed
- Chang YN, Yang JM, Bao H, Ayittey DM, Sun Z, et al. 2026. A plasma membrane-located lysophosphatidic acid acyltransferase in microalga Myrmecia incisa prefers arachidonic acid-CoA to produce glycerolipids. Appl Biochem Biotechnol. 198: 2427–2447. ArticlePubMedPDF
- Danielli M, Perne L, Jarc Jovičić E, Petan T. 2023. Lipid droplets and polyunsaturated fatty acid trafficking: Balancing life and death. Front Cell Dev Biol. 11: 1104725.ArticlePubMedPMC
- De Carvalho CC, Caramujo MJ. 2018. The various roles of fatty acids. Molecules. 23: 2583.ArticlePubMedPMC
- Falkowski PG, Raven JA. 2007. Aquatic photosynthesis, 2nd edn. Princeton University PressArticle
- FAO, Food and Agriculture Organization of the United Nations. 2020. The state of world fisheries and aquaculture 2020: Sustainability in action. Rome. Available from https://doi.org/10.4060/ca9229enArticle
- Glencross BD, Bachis E, Betancor MB, Calder P, Liland N, et al. 2025. Omega-3 futures in aquaculture: Exploring the supply and demands for long-chain omega-3 essential fatty acids by aquaculture species. Rev Fish Sci Aquac. 33: 167–216. Article
- Goo YH, Son SH, Kreienberg PB, Paul A. 2014. Novel lipid droplet-associated serine hydrolase regulates macrophage cholesterol mobilization. Arterioscler Thromb Vasc Biol. 34: 386–396. ArticlePubMedPMC
- Goo YH, Son SH, Paul A. 2017. Lipid droplet-associated hydrolase promotes lipid droplet fusion and enhances ATGL degradation and triglyceride accumulation. Sci Rep. 7: 2743.ArticlePubMedPMCPDF
- Goold H, Beisson F, Peltier G, Li-Beisson Y. 2015. Microalgal lipid droplets: Composition, diversity, biogenesis and functions. Plant Cell Rep. 34: 545–555. ArticlePubMedPDF
- Gorjão R, Azevedo-Martins AK, Rodrigues HG, Abdulkader F, Arcisio-Miranda M, et al. 2009. Comparative effects of DHA and EPA on cell function. Pharmacol Ther. 122: 56–64. ArticlePubMed
- Guiry MD, Guiry GM. 2026. AlgaeBase. Available from https://www.algaebase.orgLink
- Hu R, Xu J, Hua Y, Li Y, Li J. 2024. Could early life DHA supplementation benefit neurodevelopment? A systematic review and meta-analysis. Front Neurol. 15: 1295788.ArticlePubMedPMC
- Jaeckisch N, Yang I, Wohlrab S, Glöckner G, Kroymann J, et al. 2011. Comparative genomic and transcriptomic characterization of the toxigenic marine dinoflagellate Alexandrium ostenfeldii. PLoS One. 6: e28012. ArticlePubMedPMC
- Jiang PL, Pasaribu B, Chen CS. 2014. Nitrogen-deprivation elevates lipid levels in Symbiodinium spp. by lipid droplet accumulation: Morphological and compositional analyses. PLoS One. 9: e87416. ArticlePubMedPMC
- Kalinina V, Palii O, Safonov P, Skarlato S. 2025. Transcriptomic response of marine dinoflagellate Prorocentrum cordatum to phosphorus deficiency. Sci Rep. 15: 2014.ArticlePubMedPMCPDF
- Kang S, Kim KH, Shin HH, Kim JH, Kim BH, et al. 2025. Nitrogen excess induces cytokinesis arrest and alters lipid metabolism in the bloom-forming desmid Cosmarium tinctum. Water Res X. 28: 100394.Article
- Kim HS, Bui QTN, Wang H, Ki JS. 2023. Molecular cloning, origin, and expression of saxitoxin biosynthesis gene sxtB from the toxic dinoflagellate Alexandrium catenella. J Appl Phycol. 35: 673–685. ArticlePDF
- Kim H, Park H, Wang H, Yoo HY, Park J, et al. 2021. Low temperature and cold stress significantly increase saxitoxins (STXs) and expression of STX biosynthesis genes sxtA4 and sxtG in the dinoflagellate Alexandrium catenella. Mar Drugs. 19: 291.ArticlePubMedPMC
- Kobayashi JI, Shimbo K, Kubota T, Tsuda M. 2003. Bioactive macrolides and polyketides from marine dinoflagellates. Pure Appl Chem. 75: 337–342. Article
- Kong F, Torres-Romero I, Warakanont J, Li-Beisson Y. 2018. Lipid catabolism in microalgae: A multi-level study from genes to lipid droplets. New Phytol. 217: 134–152.Article
- Kory N, Grond S, Kamat SS, Li Z, Krahmer N, et al. 2017. Mice lacking lipid droplet-associated hydrolase, a gene linked to human prostate cancer, have normal cholesterol ester metabolism. J Lipid Res. 58: 226–235. ArticlePubMedPMC
- Kusumaningtyas P, Nurbaiti S, Suantika G, Amran MB, Nurachman Z. 2017. Enhanced oil production by the tropical marine diatom Thalassiosira sp. cultivated in outdoor photobioreactors. Appl Biochem Biotechnol. 182: 1605–1618. ArticlePubMedPDF
- La Motta LM, Padula MP, Sommer B, Camp EF, Matthews JL. 2024. Diversity of lipid profiles of Symbiodiniaceae under temperature and nutrient stress. Front Protistol. 2: 1320353.Article
- Leblond JD, Chapman PJ. 2000. Lipid class distribution of highly unsaturated long chain fatty acids in marine dinoflagellates. J Phycol. 36: 1103–1108. ArticleLink
- Lepage G, Roy CC. 1986. Direct transesterification of all classes of lipids in a one-step reaction. J Lipid Res. 27: 114–120. ArticlePubMed
- Ma XN, Chen TP, Yang B, Liu J, Chen F. 2016. Lipid production from Nannochloropsis. Mar Drugs. 14: 61.ArticlePubMedPMC
- Mendoza-Flores A, Sánchez-Saavedra MP. 2023. Light irradiance modifies the fatty acid composition of Amphidinium carterae (Dinophyceae). Phycologia. 62: 525–531. Article
- Molina-Miras A, Bueso-Sánchez A, Cerón-García MDC, Sánchez-Mirón A, Contreras-Gómez A, et al. 2022. Effect of nitrogen, phosphorous, and light colimitation on amphidinol production and growth in the marine dinoflagellate microalga Amphidinium carterae. Toxins. 14: 594.ArticlePubMedPMC
- Mooney BD, Nichols PD, De Salas MF, Hallegraeff GM. 2007. Lipid, fatty acid, and sterol composition of eight species of Kareniaceae (Dinophyta): Chemotaxonomy and putative lipid phycotoxins. J Phycol. 43: 101–111.Article
- Muhammad BL, Bui QTN, Kim HS, Ki JS. 2026. Transcriptomic insights into polyketides and toxin biosynthesis genes in freshwater dinoflagellates. Sci Rep. 6: 9472.ArticlePubMedPMCPDF
- Muhammad BL, Kim HS, Bui QTN, Ki JS. 2025. Transcriptomic comparison unveils saxitoxin biosynthesis genes in the marine dinoflagellate Gymnodinium catenatum. Harmful Algae. 147: 102872.ArticlePubMed
- Orefice I, Balzano S, Romano G, Sardo A. 2023. Amphidinium spp. as a source of antimicrobial, antifungal, and anticancer compounds. Life (Basel). 13: 2164.Article
- Qiao H, Cong C, Sun C, Li B, Wang J, et al. 2016. Effect of culture conditions on growth, fatty acid composition and DHA/EPA ratio of Phaeodactylum tricornutum. Aquaculture. 452: 311–317. Article
- Rein KS, Colon R, Romagosa CR, Ohnikian NR, Francis KT, et al. 2025. Comparative quantitative proteomic analysis of high and low toxin-producing Karenia brevis strains reveals differences in polyketide synthase abundance and redox status of the proteome. Mar Drugs. 23: 291.ArticlePubMedPMC
- Remize M, Planchon F, Loh AN, Le Grand F, Lambert C, et al. 2020. Identification of polyunsaturated fatty acids synthesis pathways in the toxic dinophyte Alexandrium minutum using 13C-labelling. Biomolecules. 10: 1428.ArticlePubMedPMC
- Research and Markets. 2021. Omega-3 market size, share & trends analysis report by type (DHA, EPA, ALA), by source (marine, plant), by application, by region, and segment forecasts, 2023-2030. Available from https://www.researchandmarkets.com/reports/4761207/omega-3-market-size-share-and-trends-analysisLink
- Řezanka T, Lukavský J, Nedbalová L, Sigler K. 2017. Lipidomic profile in three species of dinoflagellates (Amphidinium carterae, Cystodinium sp., and Peridinium aciculiferum) containing very long chain polyunsaturated fatty acids. Phytochem. 139: 88–97. ArticlePubMed
- Russo GL, Langellotti AL, Oliviero M, Sacchi R, Masi P. 2021. Sustainable production of food grade omega-3 oil using aquatic protists: Reliability and future horizons. New Biotechnol. 62: 32–39. ArticlePubMed
- Ryckebosch E, Bruneel C, Muylaert K, Foubert I. 2012. Microalgae as an alternative source of omega-3 long chain polyunsaturated fatty acids. Lipid Technol. 24: 128–130. Article
- Ryther JH, Guillard RRL. 1962. Studies of marine planktonic diatoms: II. Use of Cyclotella nana Hustedt for assays of vitamin B12 in sea water. Can J Microbiol. 8: 437–445. Article
- Sampayo EM, Franceschinis L, Hoegh-Guldberg O, Dove S. 2007. Niche partitioning of closely related symbiotic dinoflagellates. Mol Ecol. 16: 3721–3733. ArticlePubMed
- Schmitz C, Sforca ML, Maraschin M. 2025. Metabolomic analysis and computational biology reveal the potential metabolic pathways for improvement of fucoxanthin and eicosapentaenoic acid (EPA) biosynthesis in Phaeodactylum tricornutum. Appl Biochem Biotechnol. 197: 3132–3151. ArticlePubMedPDF
- Singh J, Saxena RC. 2015. An introduction to microalgae: Diversity and significance. In Kim SK. (ed.), Handbook of marine microalgae: Biotechnology advances, pp. 11–24. Academic PressArticle
- Stephens TG, Ragan MA, Bhattacharya D, Chan CX. 2021. Core genes in the fatty acid synthesis pathway of alveolates. Genome Biol Evol. 13: evab242.ArticlePubMedPMC
- Suh SS, Kim SJ, Hwang J, Park M, Lee TK, et al. 2015. Fatty acid methyl ester profiles and nutritive values of 20 marine microalgae in Korea. Asian Pac J Trop Med. 8: 191–196. ArticlePubMed
- Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 30: 2725–2729. ArticlePubMedPMC
- Taylor FJR, Hoppenrath M, Saldarriaga JF. 2008. Dinoflagellate diversity and distribution. Biodivers Conserv. 17: 407–418. ArticlePDF
- Thiyagarajan S, Arumugam M, Kathiresan S. 2020. Identification and functional characterization of two novel fatty acid genes from marine microalgae for eicosapentaenoic acid production. Appl Biochem Biotechnol. 190: 1371–1384. ArticlePubMedPDF
- Tocher DR, Betancor MB, Sprague M, Olsen RE, Napier JA. 2019. Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: Bridging the gap between supply and demand. Nutrients. 11: 89.ArticlePubMedPMC
- Uauy R, Dangour AD. 2006. Nutrition in brain development and aging: Role of essential fatty acids. Nutr Rev. 64: S24–S33. ArticlePubMed
- Vazhappilly R, Chen F. 1998. Eicosapentaenoic acid and docosahexaenoic acid production potential of microalgae and their heterotrophic growth. J Am Oil Chem Soc. 75: 393–397. ArticleLink
- Volkman JK, Jeffrey SW, Nichols PD, Rogers GI, Garland CD. 1989. Fatty acid and lipid composition of 10 species of microalgae used in mariculture. J Exp Mar Biol Ecol. 128: 219–240. Article
- Wang H, Kim H, Ki JS. 2021a. Preliminary result of de novo transcriptome sequencing of the marine toxic dinoflagellate Alexandrium catenella incubated under several different stresses. Mar Biol. 168: 104.ArticlePDF
- Wang H, Kim H, Ki JS. 2021b. Transcriptome survey, molecular identification, and expression analysis of stress-responsive genes in the toxic dinoflagellate Alexandrium pacificum under algicidal agents and metal stresses. J Appl Phycol. 33: 3139–3151. ArticlePDF
- Weng LC, Pasaribu B, Lin IP, Tsai CH, Chen CS, et al. 2014. Nitrogen deprivation induces lipid droplet accumulation and alters fatty acid metabolism in symbiotic dinoflagellates isolated from Aiptasia pulchella. Sci Rep. 4: 5777.ArticlePubMedPMCPDF
- Wilfling F, Haas JT, Walther TC, Farese RV Jr. 2014. Lipid droplet biogenesis. Curr Opin Cell Biol. 29: 39–45. ArticlePubMedPMC
- Yin FW, Zhan CT, Huang J, Sun XL, Yin LF, et al. 2023. Efficient co-production of docosahexaenoic acid oil and carotenoids in Aurantiochytrium sp. using a light intensity gradient strategy. Appl Biochem Biotechnol. 195: 623–638. ArticlePubMedPDF
- Yoneda A, Ishibashi Y, Yoshida M, Watanabe MM, Ito M, et al. 2022. Proteomic and lipidomic analyses of lipid droplets in Aurantiochytrium limacinum ATCC MYA-1381. Algal Res. 67: 102844.Article
- Zhao Y, Cao R, Li J, Xu Y, Zhou L, et al. 2025. Lipid droplets in plants: Turnover and stress responses. Front Plant Sci. 16: 1625830.ArticlePubMedPMC
Citations
Citations to this article as recorded by




 ePub Link
ePub Link Cite this Article
Cite this Article