- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- For Contributors
- Policies
- E-Submission

- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- Policies
- For Contributors
Articles
- Page Path
- HOME > J. Microbiol > Volume 63(5); 2025 > Article
-
Full article
Fungal diversity from Fildes Peninsula (Antarctica) and their antibiosis bioactivity against two plant pathogens - Ji Seon Kim1, Enzo Romero2, Yoonhee Cho1, Ramón Ahumada-Rudolph2, Christian Núñez2, Jonhatan Gómez-Espinoza3, Ernesto Moya-Elizondo4, Sigisfredo Garnica5, Young Woon Lim1,*, Jaime R. Cabrera-Pardo2,6,*
-
Journal of Microbiology 2025;63(5):e2411029.
DOI: https://doi.org/10.71150/jm.2411029
Published online: April 14, 2025
1School of Biological Sciences and Institute of Biodiversity, Seoul National University, Seoul 08826, Republic of Korea
2Applied and Sustainable Chemistry Laboratory (LabQAS), Department of Chemistry, Universidad del Bío-Bío, Concepción 4051381, Chile
3Technical Professional High School Diego Portales, Department of Science, Linares, Chile
4Department of Plant Production, Faculty of Agronomy, Universidad de Concepción, Concepción 3812120, Chile
5Institute of Biochemistry and Microbiology, Faculty of Sciences, Universidad Austral de Chile, Teja Island, Valdivia 5090000, Chile
6College of Dental Medicine, Roseman University of Health Sciences, South Jordan, UT 84095, USA
- *Correspondence Young Woon Lim ywlim@snu.ac.kr Jaime R. Cabrera-Pardo jacabrera@ubiobio.cl
© The Author(s), under exclusive licence to Microbiological Society of Korea 2026
This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (CC BY 4.0) (https://creativecommons.org/licenses/by/4.0/) which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
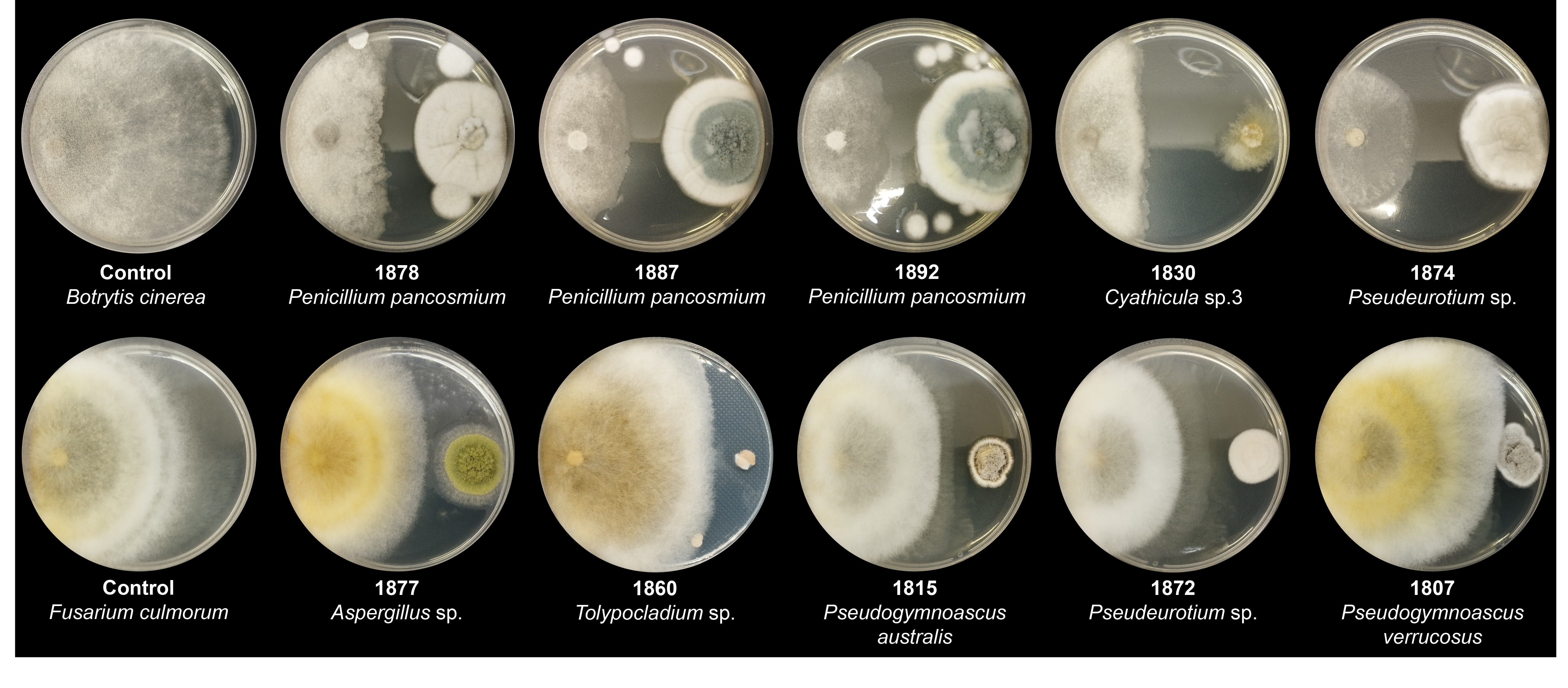
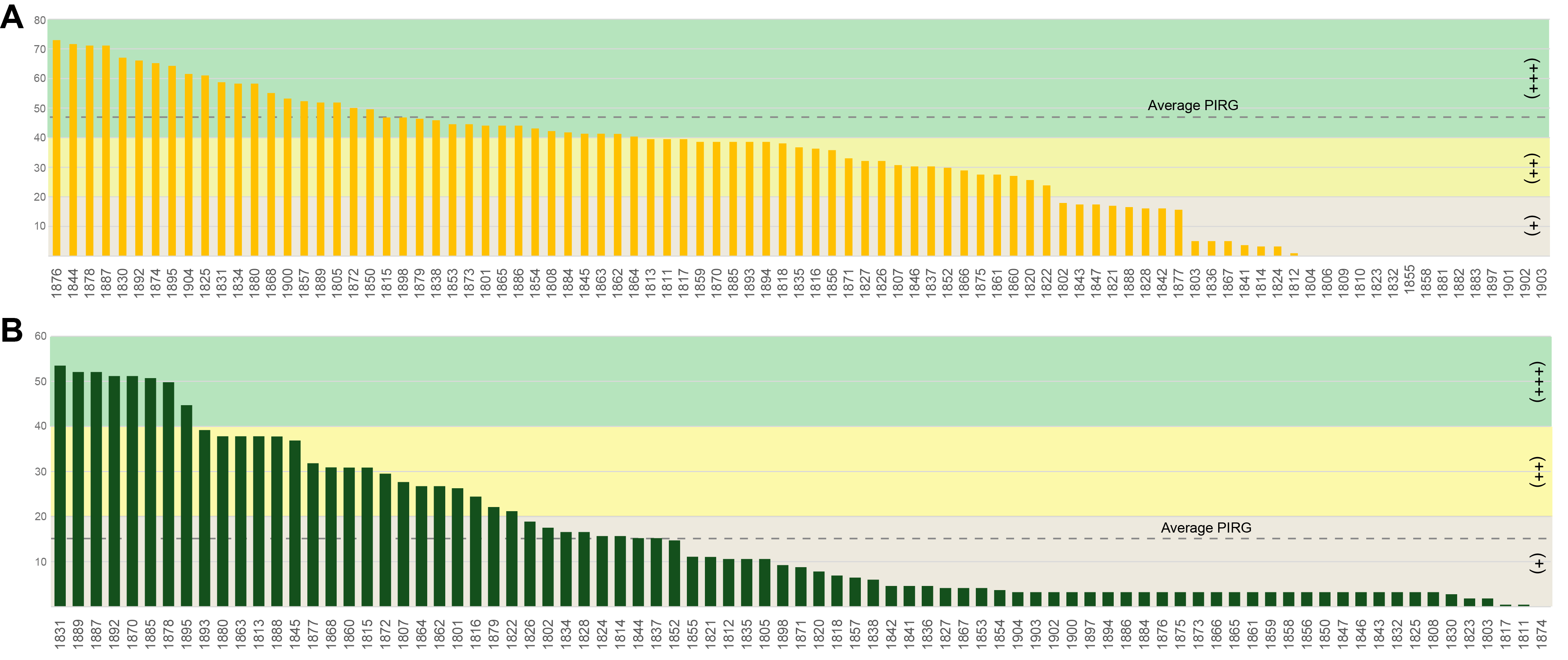
- Antarctic fungi can effectively adapt to extreme environments, which leads to the production of unique bioactive compounds. Studies on the discovery of fungi in the diverse environments of Antarctica and their potential applications are increasing, yet remain limited. In this study, fungi were isolated from various substrates on the Fildes Peninsula in Antarctica and screened for their antibiosis activity against two significant plant pathogenic fungi, Botrytis cinerea and Fusarium culmorum. Phylogenetic analysis using multiple genetic markers revealed that the isolated Antarctic fungal strains are diverse, some of which are novel, emphasizing the underexplored biodiversity of Antarctic fungi. These findings suggest that these fungi have potential for the development of new antifungal agents that can be applied in agriculture to manage fungal plant pathogens. Furthermore, the antibiosis activities of the isolated Antarctic fungi were evaluated using a dual-culture assay. The results indicated that several strains from the genera Cyathicula, Penicillium, and Pseudeurotium significantly inhibited pathogen growth, with Penicillium pancosmium showing the highest inhibitory activity against Botrytis cinerea. Similarly, Aspergillus and Tolypocladium strains exhibited strong antagonistic effects against Fusarium culmorum. This study enhances our understanding of Antarctic fungal diversity and highlights its potential for biotechnological applications.
Introduction
Materials and Methods
Results
Discussion
Acknowledgments
This study was supported by the Instituto Antártico Chileno (INACH) (grant number: INACH RT_16-21), VRIP-Universidad del Bío-Bío (grant numbers: GI2310643 and EQ2326450), and the Korea Institute of Marine Science & Technology Promotion (KIMST) funded by the Ministry of Oceans and Fisheries (grant number: RS-2024-00341152).
Conflict of interest
None.
Ethical Statements
Not applicable.
Supplementary Information
Fig. S1.
- Aly AH, Debbab A, Proksch P. 2011. Fifty years of drug discovery from fungi. Fungal Divers. 50: 3–19. ArticlePDF
- Antipova TV, Zaitsev KV, Zhelifonova VP, Tarlachkov SV, Grishin YK, et al. 2023. The potential of Arctic Pseudogymnoascus fungi in the biosynthesis of natural products. Fermentation. 9(8): 702.Article
- Bar-On YM, Phillips R, Milo R. 2018. The biomass distribution on Earth. Proc Natl Acad Sci USA. 115(25): 6506–6511. ArticlePubMedPMC
- Bates ST, Berg-Lyons D, Lauber CL, Walters WA, Knight R, et al. 2012. A preliminary survey of lichen-associated eukaryotes using pyrosequencing. Lichenologist. 44(1): 137–146. Article
- Bensch K, Braun U, Groenewald JZ, Crous PW. 2012. The genus Cladosporium. Stud Mycol. 72: 1–401. ArticlePubMedPMC
- Bhattarai K, Bhattarai K, Kabir ME, Bastola R, Baral B. 2021. Fungal natural products galaxy: Biochemistry and molecular genetics toward blockbuster drugs discovery. Adv Genet. 107: 193–284. ArticlePubMed
- Bladt T, Frisvad J, Knudsen P, Larsen T. 2013. Anticancer and antifungal compounds from Aspergillus, Penicillium, and other filamentous fungi. Molecules. 18(9): 11338–11376. ArticlePubMedPMC
- Brown AG, Smale TC, King TJ, Hasenkamp R, Thompson RH. 1976. Crystal and molecular structure of compactin, a new antifungal metabolite from Penicillium brevicompactum. J Chem Soc Perkin Trans 1. 11: 1165–1170. Article
- Bushley KE, Raja R, Jaiswal P, Cumbie JS, Nonogaki M, et al. 2013. The genome of Tolypocladium inflatum: Evolution, organization, and expression of the cyclosporin biosynthetic gene cluster. PLoS Genet. 9(6): e1003496. ArticlePubMedPMC
- Câmara PEAS, Convey P, Rangel SB, Konrath M, Barreto CC, et al. 2021. The largest moss carpet transplant in Antarctica and its bryosphere cryptic biodiversity. Extremophiles. 25(4): 369–384. ArticlePubMedPDF
- Chen L, Yue Q, Zhang X, Xiang M, Wang C, et al. 2013. Genomics-driven discovery of the pneumocandin biosynthetic gene cluster in the fungus Glarea lozoyensis. BMC Genomics. 14(1): 339.ArticlePubMedPMCPDF
- De Carvalho CR, Santiago IF, Da Costa Coelho L, Câmara PEAS, Silva MC, et al. 2019. Fungi associated with plants and lichens of Antarctica, In Rosa LH. (ed.), Fungi of Antarctica, pp. 165–199, Springer.
- Dean R, Van Kan JAL, Pretorius ZA, Hammond‐Kosack KE, Di Pietro A, et al. 2012. The top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol. 13(4): 414–430. ArticlePubMedPMC
- Dupuis JR, Roe AD, Sperling FAH. 2012. Multi-locus species delimitation in closely related animals and fungi: One marker is not enough. Mol Ecol. 21(18): 4422–4436. ArticlePubMed
- Dwivedi M, Singh P, Pandey AK. 2024. Botrytis fruit rot management: What have we achieved so far? Food Microbiol. 122: 104564.ArticlePubMed
- Elhamouly NA, Hewedy OA, Zaitoon A, Miraples A, Elshorbagy OT, et al. 2022. The hidden power of secondary metabolites in plant-fungi interactions and sustainable phytoremediation. Front Plant Sci. 13: 1044896.ArticlePubMedPMC
- Gholami-Shabani M, Shams-Ghahfarokhi M, Razzaghi-Abyaneh M. 2019. Natural product synthesis by fungi: Recent trends and future prospects. In Yadav AN, Singh S, Mishra S, Gupta A. (eds.), Recent advancement in white biotechnology through fungi, pp. 195–228, Springer.
- González J, Romero-Aguilar L, Matus-Ortega G, Pablo Pardo J, Flores-Alanis A, et al. 2020. Levaduras adaptadas al frío: El tesoro biotecnológico de la Antártica. TIP Rev Esp Cienc Quím Biol. 23.
- Gryganskyi AP, Humber RA, Smith ME, Miadlikovska J, Wu S, et al. 2012. Molecular phylogeny of the Entomophthoromycota. Mol Phylogenet Evol. 65(2): 682–694. ArticlePubMed
- Hahn M. 2014. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J Chem Biol. 7(4): 133–141. ArticlePubMedPMCPDF
- Hassan N, Rafiq M, Hayat M, Shah AA, Hasan F. 2016. Psychrophilic and psychrotrophic fungi: A comprehensive review. Rev Environ Sci Biotechnol. 15(2): 147–172. ArticlePDF
- Henríquez M, Vergara K, Norambuena J, Beiza A, Maza F, et al. 2014. Diversity of cultivable fungi associated with Antarctic marine sponges and screening for their antimicrobial, antitumoral and antioxidant potential. World J Microbiol Biotechnol. 30(1): 65–76. ArticlePubMedPDF
- Heo YM, Lee H, Kim K, Kwon SL, Park MY, et al. 2019. Fungal diversity in intertidal mudflats and abandoned solar salterns as a source for biological resources. Mar Drugs. 17(11): 601.ArticlePubMedPMC
- Herrera-Défaz M, Fuentealba D, Dibona-Villanueva L, Schwantes D, Jiménez B, et al. 2023. Biocontrol of Botrytis cinerea on grape berries in Chile: Use of registered biofungicides and a new chitosan-based fungicide. Horticulturae. 9(7): 746.Article
- Hirose D, Hobara S, Matsuoka S, Kato K, Tanabe Y, et al. 2016. Diversity and community assembly of moss-associated fungi in ice-free coastal outcrops of continental Antarctica. Fungal Ecol. 24: 94–101. Article
- Hirose D, Hobara S, Tanabe Y, Uchida M, Kudoh S, et al. 2017. Abundance, richness, and succession of microfungi in relation to chemical changes in Antarctic moss profiles. Polar Biol. 40(12): 2457–2468. ArticlePDF
- Ismail MA, Amin MA, Eid AM, Hassan SED, Mahgoub HAM, et al. 2021. Comparative study between exogenously applied plant growth hormones versus metabolites of microbial endophytes as plant growth-promoting for Phaseolus vulgaris L. Cells. 10(5): 1059.ArticlePubMedPMC
- Johnston PR, Quijada L, Smith CA, Baral HO, Hosoya T, et al. 2019. A multigene phylogeny toward a new phylogenetic classification of Leotiomycetes. IMA Fungus. 10(1): 1.ArticlePubMedPMCPDF
- Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, et al. 2012. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 28(12): 1647–1649. ArticlePubMedPMCPDF
- Kendrick B. 2011. Fungi: Ecological importance and impact on humans. In Encyclopedia of Life Sciences. Wiley.
- Khokhar I, Mukhtar I, Mushtaq S. 2011. Antifungal effect of Penicillium metabolites against some fungi. Arch Phytopathol Plant Prot. 44(14): 1347–1351. Article
- Kim BS, Hwang BK. 2007. Microbial fungicides in the control of plant diseases. J Phytopathol. 155(11-12): 641–653. Article
- Kiss L. 2012. Limits of nuclear ribosomal DNA internal transcribed spacer (ITS) sequences as species barcodes for fungi. Proc Natl Acad Sci USA. 109(27): E1811.ArticlePubMedPMC
- Kochkina GA, Ivanushkina NE, Lupachev AV, Starodumova IP, Vasilenko OV, et al. 2019. Diversity of mycelial fungi in natural and human-affected Antarctic soils. Polar Biol. 42(1): 47–64. ArticlePDF
- Kochkina GA, Ozerskaya SM, Ivanushkina NE, Chigineva NI, Vasilenko OV, et al. 2014. Fungal diversity in the Antarctic active layer. Microbiology. 83(1–2): 94–101. ArticlePDF
- Latorre BA, Elfar K, Ferrada EE. 2015. Gray mold caused by Botrytis cinerea limits grape production in Chile. Cienc Inv Agrar. 42(3): 305–330. Article
- Marx JC, Collins T, D’Amico S, Feller G, Gerday C. 2007. Cold-adapted enzymes from marine Antarctic microorganisms. Mar Biotechnol. 9: 293–304. ArticlePDF
- Mishra PK, Fox RTV, Culham A. 2000. Application of nr‐DNA ITS sequence for identification of Fusarium culmorum isolates. EPPO Bull. 30(3–4): 493–498. Article
- Naranjo‐Ortiz MA, Gabaldón T. 2019. Fungal evolution: Major ecological adaptations and evolutionary transitions. Biol Rev. 94(4): 1443–1476. ArticlePubMedPMCLink
- Núñez-Montero K, Barrientos L. 2018. Advances in Antarctic research for antimicrobial discovery: A comprehensive narrative review of bacteria from Antarctic environments as potential sources of novel antibiotic compounds against human pathogens and microorganisms of industrial importance. Antibiotics. 7(4): 90.ArticlePubMedPMC
- O’Donnell K, Nirenberg HI, Aoki T, Cigelnik E. 2000. A multigene phylogeny of the Gibberella fujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience. 41(1): 61–78. Article
- O’Donnell K, Sutton DA, Fothergill A, McCarthy D, Rinaldi MG, et al. 2008. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium solani species complex. J Clin Microbiol. 46(8): 2477–2490. ArticlePubMedPMCLink
- Ordóñez-Enireb E, Cucalón RV, Cárdenas D, Ordóñez N, Coello S, et al. 2022. Antarctic fungi with antibiotic potential isolated from Fort William Point, Antarctica. Sci Rep. 12(1): 21477.PubMedPMC
- Park CH, Kim KM, Elvebakk A, Kim O, Jeong G, et al. 2015. Algal and fungal diversity in Antarctic lichens. J Eukaryot Microbiol. 62(2): 196–205. ArticlePubMedLink
- Price MN, Dehal PS, Arkin AP. 2010. FastTree 2-Approximately maximum-likelihood trees for large alignments. PLoS One. 5(3): e9490. ArticlePubMedPMC
- Quandt CA, Bushley KE, Spatafora JW. 2015. The genome of the truffle-parasite Tolypocladium ophioglossoides and the evolution of antifungal peptaibiotics. BMC Genomics. 16(1): 553.ArticlePubMedPMCPDF
- Quandt CA, Haelewaters D. 2021. Phylogenetic advances in Leotiomycetes, an understudied clade of taxonomically and ecologically diverse fungi. In Encyclopedia of Mycology, pp. 284–294, Elsevier.
- Rämä T, Hassett BT, Bubnova E. 2017. Arctic marine fungi: From filaments and flagella to operational taxonomic units and beyond. Bot Mar. 60(4): 433–452.
- Ramasamy KP, Mahawar L, Rajasabapathy R, Rajeshwari K, Miceli C, et al. 2023. Comprehensive insights on environmental adaptation strategies in Antarctic bacteria and biotechnological applications of cold-adapted molecules. Front Microbiol. 14: 1197797.ArticlePubMedPMC
- Rastegari AA, Yadav AN, Yadav N. 2020. New and Future Developments in Microbial Biotechnology and Bioengineering: Trends of Microbial Biotechnology for Sustainable Agriculture and Biomedicine Systems: Diversity and Functional Perspectives, Elsevier.
- Roca-Couso R, Flores-Félix JD, Rivas R. 2021. Mechanisms of action of microbial biocontrol agents against Botrytis cinerea. J Fungi. 7(12): 1045.ArticlePubMedPMC
- Rosa LH, Da Costa Coelho L, Pinto OHB, Carvalho-Silva M, Convey P, et al. 2021. Ecological succession of fungal and bacterial communities in Antarctic mosses affected by a fairy ring disease. Extremophiles. 25(5–6): 471–481. ArticlePubMedPDF
- Rosa LH, De Sousa JRP, De Menezes GCA, Da Costa Coelho L, Carvalho-Silva M, et al. 2020. Opportunistic fungi found in fairy rings are present on different moss species in the Antarctic Peninsula. Polar Biol. 43(5): 587–596. ArticlePDF
- Rosa LH, Zani CL, Cantrell CL, Duke SO, Van Dijck P, et al. 2019. Fungi in Antarctica: Diversity, ecology, effects of climate change, and bioprospection for bioactive compounds. In Rosa LH. (ed.), Fungi of Antarctica, pp. 1–17, Springer.
- Scherm B, Balmas V, Spanu F, Pani G, Delogu G, et al. 2013. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol Plant Pathol. 14(4): 323–341. PubMed
- Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, et al. 2012. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc Natl Acad Sci USA. 109(16): 6241–6246. PubMedPMC
- Schueffler A, Anke T. 2014. Fungal natural products in research and development. Nat Prod Rep. 31(10): 1425–1448. ArticlePubMed
- Shi T, Li XQ, Wang ZM, Zheng L, Yu YY, et al. 2022. Bioactivity-guided screening of antimicrobial secondary metabolites from Antarctic cultivable fungus Acrostalagmus luteoalbus CH-6 combined with molecular networking. Mar Drugs. 20(5): 334.ArticlePubMedPMC
- Shi T, Li XQ, Zheng L, Zhang YH, Dai JJ, et al. 2021. Sesquiterpenoids from the Antarctic fungus Pseudogymnoascus sp. HSX2#-11. Front Microbiol. 12: 688202.ArticlePubMedPMC
- Stamatakis A. 2014. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 30(9): 1312–1313. ArticlePubMedPMCPDF
- Suarez MB, Walsh K, Boonham N, O’Neill T, Pearson S, et al. 2005. Development of real-time PCR (TaqMan®) assays for the detection and quantification of Botrytis cinerea in planta. Plant Physiol Biochem. 43(9): 890–899. ArticlePubMed
- Thambugala KM, Daranagama DA, Phillips AJL, Kannangara SD, Promputtha I, et al. 2020. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front Cell Infect Microbiol. 10: 604923.ArticlePubMedPMC
- Varrella S, Barone G, Tangherlini M, Rastelli E, Dell’Anno A, et al. 2021. Diversity, ecological role and biotechnological potential of Antarctic marine fungi. J Fungi. 7(5): 391.ArticlePubMedPMC
- Vicente MF, Basilio A, Cabello A, Peláez F. 2003. Microbial natural products as a source of antifungals. Clin Microbiol Infect. 9(1): 15–32. ArticlePubMed
- Vieira G, Purić J, Morão LG, Dos Santos JA, Inforsato FJ, et al. 2018. Terrestrial and marine Antarctic fungi extracts active against Xanthomonas citri subsp. citri. Lett Appl Microbiol. 67(1): 64–71. ArticlePubMedLink
- Vinale F, Sivasithamparam K, Ghisalberti EL, Woo SL, Nigro M, et al. 2014. Trichoderma secondary metabolites active on plants and fungal pathogens. Open Mycol J. 8(1): 127–139. Article
- Visagie CM, Houbraken J, Frisvad JC, Hong SB, Klaassen CHW, et al. 2014. Identification and nomenclature of the genus Penicillium. Stud Mycol. 78(1): 343–371. ArticlePubMedPMC
- Wang J, Liu M, Mao C, Li S, Zhou J, et al. 2023. Comparative proteomics reveals the mechanism of cyclosporine production and mycelial growth in Tolypocladium inflatum affected by different carbon sources. Front Microbiol. 14: 1259101.ArticlePubMedPMC
- Wang X, Radwan MM, Taráwneh AH, Gao J, Wedge DE, et al. 2013. Antifungal activity against plant pathogens of metabolites from the endophytic fungus Cladosporium cladosporioides. J Agric Food Chem. 61(19): 4551–4555. ArticlePubMedPMC
- White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, pp. 315–322, Elsevier.
- Yin Y, Miao J, Shao W, Liu X, Zhao Y, et al. 2023. Fungicide resistance: Progress in understanding mechanism, monitoring, and management. Phytopathology. 113(4): 707–718. ArticlePubMed
- Yu NH, Park SY, Kim JA, Park CH, Jeong MH, et al. 2018. Endophytic and endolichenic fungal diversity in maritime Antarctica based on cultured material and their evolutionary position among Dikarya. Fungal Syst Evol. 2: 263–272. ArticlePubMedPMC
- Zucconi L, Canini F, Temporiti ME, Tosi S. 2020. Extracellular enzymes and bioactive compounds from Antarctic terrestrial fungi for bioprospecting. Int J Environ Res Public Health. 17(18): 6459.ArticlePubMedPMC
References
Supplementary Information
References
Citations
- A Drought-Activated Bacterial Symbiont Enhances Legume Resilience Through Coordinated Amino Acid Metabolism
Susmita Das Nishu, Jee Hyun No, Gui Nam Wee, Tae Kwon Lee
Microorganisms.2026; 14(1): 114. CrossRef - Biosynthetic potential of endophytic fungi from tropical medicinal plants: genomic and metabolomic perspectives
Asri Peni Wulandari, Erin Nur Lestari, Ayu Wandira, Rezqita Putri Pitaloka, Laita Nurjannah, Azmi Azhari
Mycology.2026;[Epub] CrossRef - Agaricales from Antarctica: Diversity of basidiomata, research challenges, and future perspectives in polar environments
Fernando Augusto Bertazzo-Silva, Jair Putzke
Fungal Biology Reviews.2025; 54: 100458. CrossRef - Diversity, geographical distribution and environmental adaptations of snow molds
Tamotsu Hoshino
Mycoscience.2025; 66(6): 334. CrossRef
 ePub Link
ePub Link Cite this Article
Cite this Article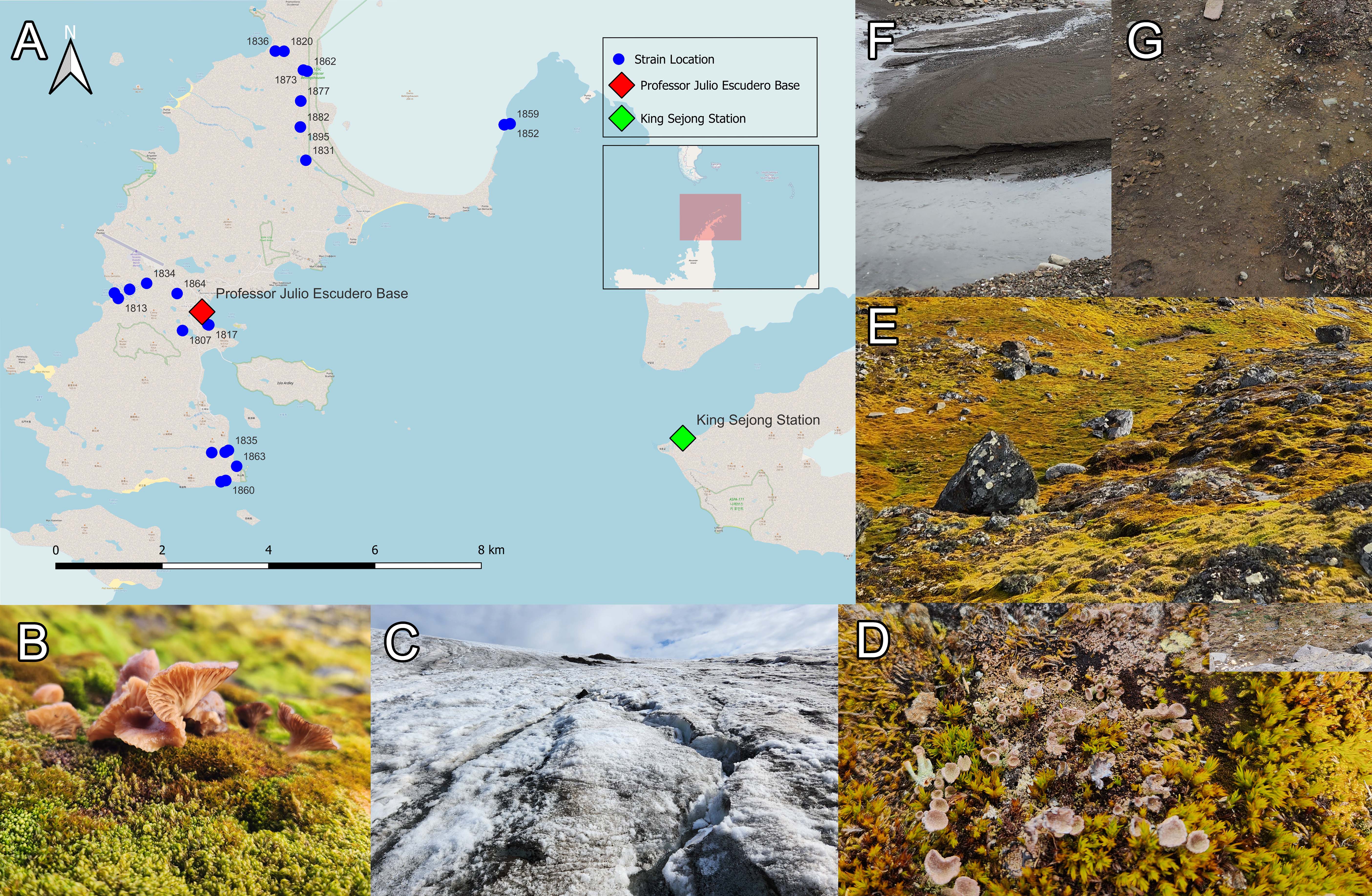

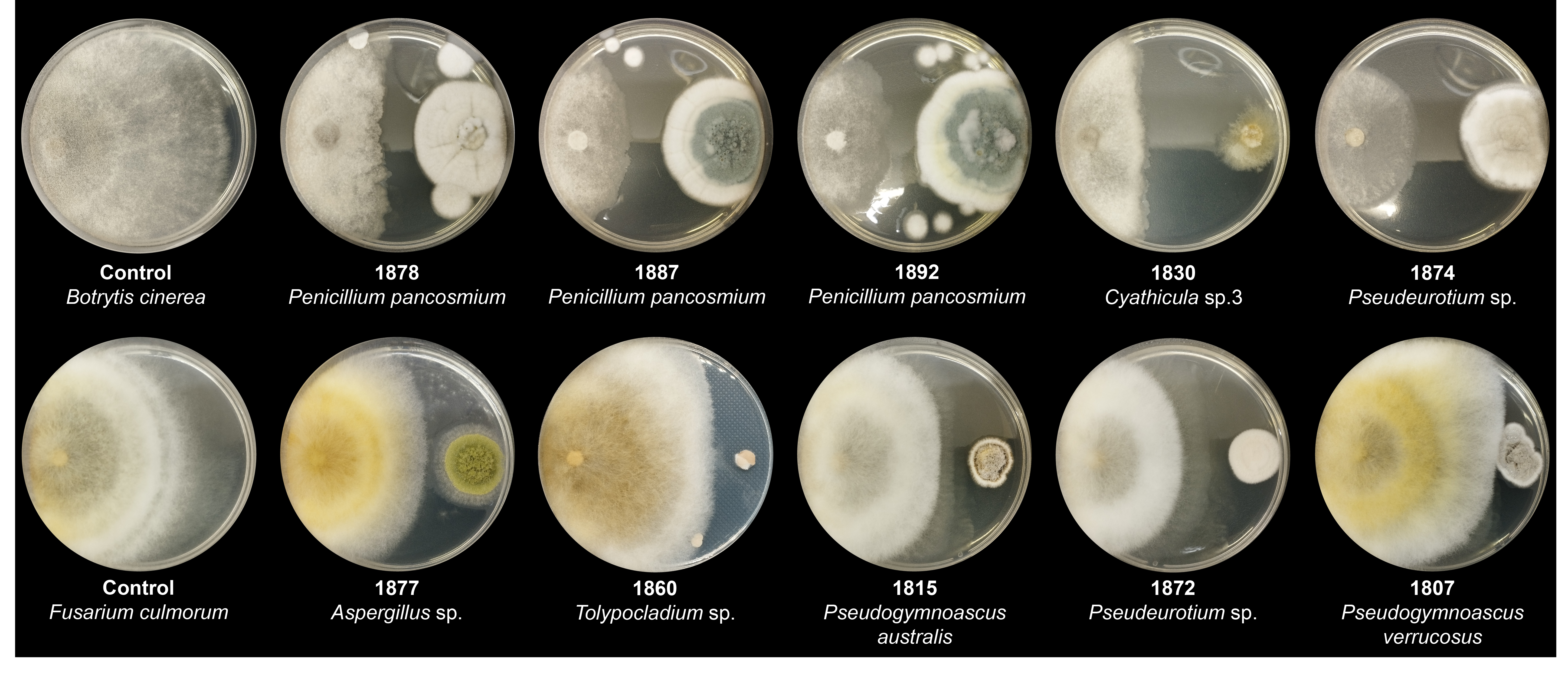
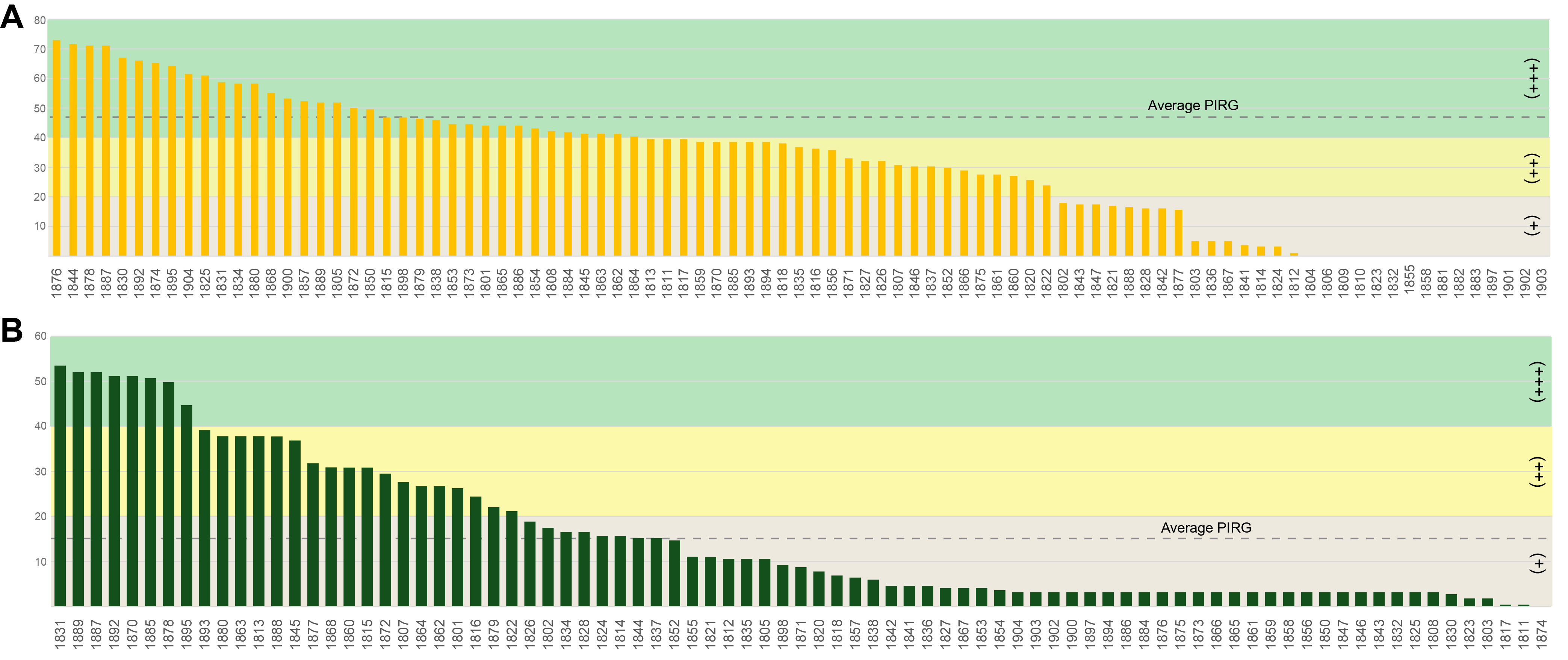




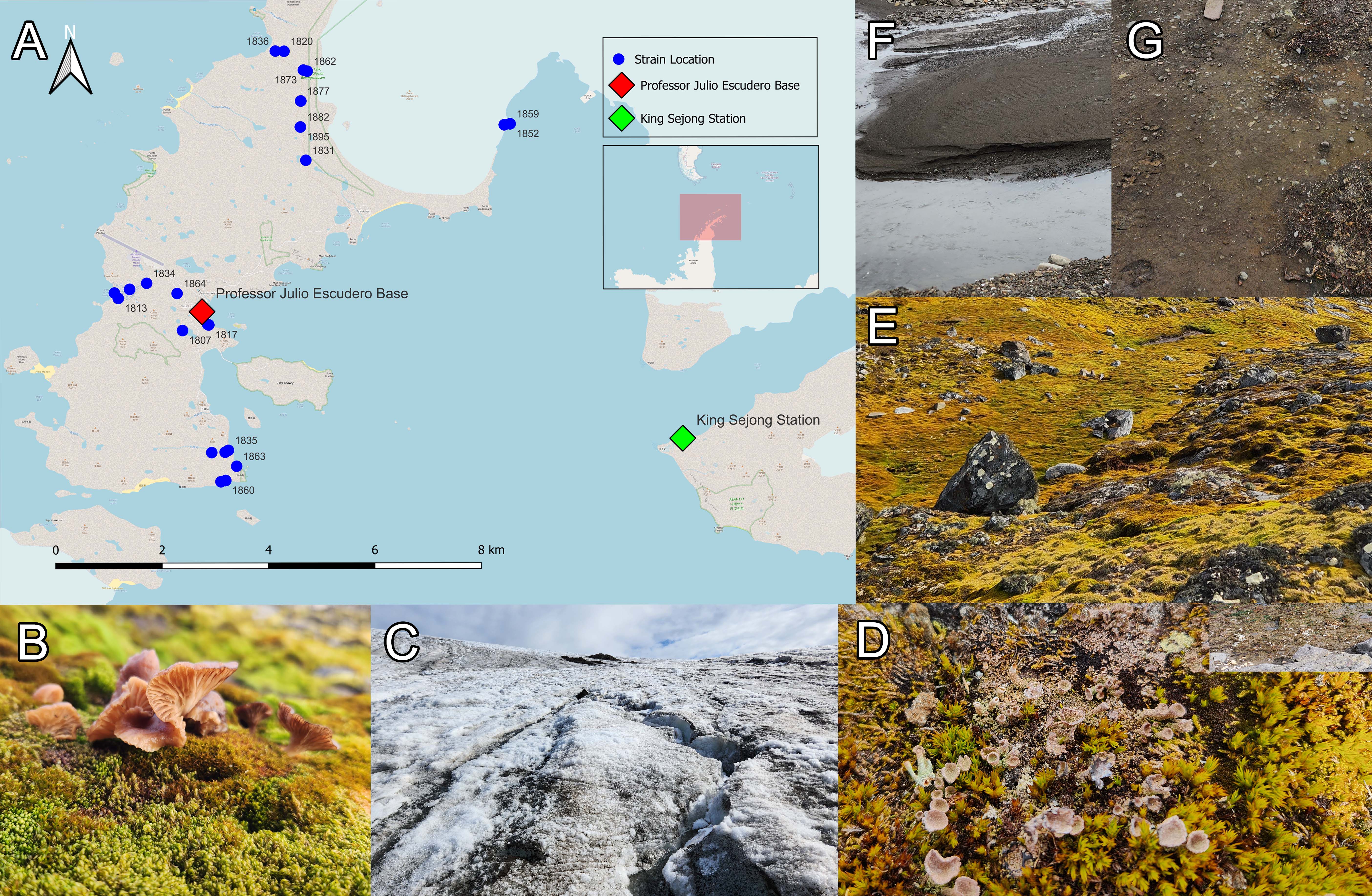
Fig. 1.
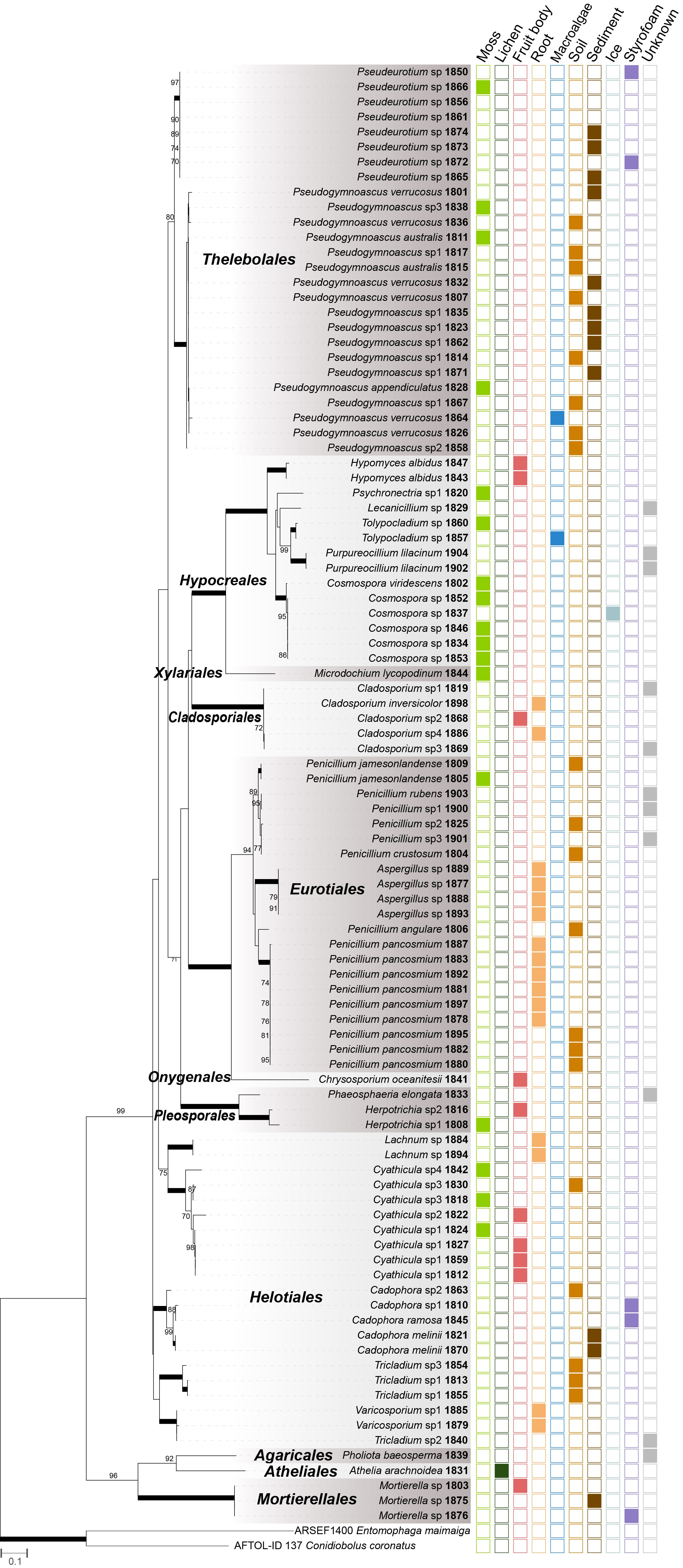
Fig. 2.
Fig. 3.
Fig. 4.
| Order | Family | Species identification | Strain NUM | Substrate | Latitude (S) | Longitude (W) | ITS | LSU | TUB | CMD | ACT | TEF1 | RPB2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Agaricales | Strophariaceae | Pholiota baeosperma | 1839 | - | - | - | PQ427716 | PQ427669 | |||||
| Amphisphaeriales | Amphisphaeriaceae | Microdochium lycopodinum | 1844 | Moss | 62°10'11.4168" | 58°51'10.2096" | PQ427765 | PQ427672 | |||||
| Atheliales | Atheliaceae | Athelia arachnoidea | 1831 | Lichen | 62°10'32.9340" | 58°55'28.6860" | PQ427721 | ||||||
| Cladosporiales | Cladosporiaceae | Cladosporium inversicolor | 1898 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427783 | PQ433547 | |||||
| Cladosporium sp.1 | 1819 | - | - | - | PQ427699 | ||||||||
| Cladosporium sp.2 | 1868 | Fruit body | 62°10'10.8408" | 58°51'02.6460" | PQ427748 | PQ433531 | PQ433545 | ||||||
| Cladosporium sp.3 | 1869 | - | - | - | PQ427747 | ||||||||
| Cladosporium sp.4 | 1886 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427692 | PQ433530 | PQ433546 | ||||||
| Eurotiales | Aspergillaceae | Aspergillus sp. | 1877 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427757 | PQ456765 | |||||
| 1888 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427709 | |||||||||
| 1889 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427694 | PQ456764 | ||||||||
| 1893 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427710 | |||||||||
| Penicillium angulare | 1806 | Soil | 62°10'11.4168" | 58°51'10.2096" | PQ427704 | PQ456772 | |||||||
| Penicillium crustosum | 1804 | Soil | 62°13'48.2520" | 58°57'19.5336" | PQ427740 | PQ456773 | |||||||
| Penicillium jamesonlandense | 1805 | Moss | 62°10'11.4168" | 58°51'10.2096" | PQ427738 | PQ456774 | |||||||
| 1809 | Soil | 62°10'11.4168" | 58°51'10.2096" | PQ427739 | PQ456775 | ||||||||
| Penicillium pancosmium | 1878 | Root | 62°10'11.4168" | 58°51'10.2096" | PQ427742 | PQ456783 | |||||||
| 1880 | Soil | 62°10'12.7" | 58°55'35.8" | PQ427717 | PQ456780 | PQ433532 | |||||||
| 1881 | Root | 62°10'12.7" | 58°55'35.8" | PQ427707 | PQ456778 | PQ433533 | |||||||
| 1882 | Soil | 62°10'12.7" | 58°55'35.8" | PQ427697 | PQ456777 | ||||||||
| 1883 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427689 | PQ456776 | PQ433534 | |||||||
| 1887 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427737 | PQ456782 | PQ433535 | |||||||
| 1892 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427708 | PQ456779 | PQ433536 | |||||||
| 1895 | Soil | 62°10'12.7" | 58°55'35.8" | PQ427743 | PQ456784 | ||||||||
| 1897 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427736 | PQ456781 | PQ433537 | |||||||
| Penicillium rubens | 1903 | - | - | - | PQ427741 | PQ456785 | PQ433538 | ||||||
| Penicillium sp.1 | 1900 | - | - | - | PQ427696 | PQ456786 | |||||||
| Penicillium sp.2 | 1825 | Soil | 62°12'16.3656" | 58°58'09.4368" | PQ427735 | PQ456787 | |||||||
| Penicillium sp.3 | 1901 | - | - | - | PQ427773 | PQ456788 | |||||||
| Helotiales | Discinellaceae | Varicosporium sp. | 1879 | Root | 62°10'11.4168" | 58°51'10.2096" | PQ427701 | PQ427685 | |||||
| 1885 | Root | 62°10'11.4168" | 58°51'10.2096" | PQ427763 | PQ427686 | ||||||||
| Helotiaceae | Cyathicula sp.1 | 1812 | Fruit body | 62°13'47.4816" | 58°57'13.3560" | PQ427754 | |||||||
| 1824 | Moss | 62°11'51.3420" | 58°59'18.3408" | PQ427753 | |||||||||
| 1827 | Fruit body | 62°13'47.4816" | 58°57'13.3560" | PQ427732 | |||||||||
| 1859 | Fruit body | 62°10'10.8408" | 58°51'02.6460" | PQ427777 | |||||||||
| Cyathicula sp.2 | 1822 | Fruit body | 62°12'12.69" | 58°57'36.59" | PQ427760 | ||||||||
| Cyathicula sp.3 | 1818 | Moss | 62°12'12.9708" | 58°57'35.2908" | PQ427713 | ||||||||
| 1830 | Soil | 62°12'12.9708" | 58°57'35.2908" | PQ427734 | |||||||||
| Cyathicula sp.4 | 1842 | Moss | 62°09'26.6148" | 58°55'57.0360" | PQ427755 | ||||||||
| Lachnaceae | Lachnum sp. | 1884 | Root | 62°09'56.8980" | 58°55'35.2488" | PQ427781 | |||||||
| 1894 | Root | 62°10'11.4168" | 58°51'10.2096" | PQ427782 | |||||||||
| Ploettnerulaceae | Cadophora melinii | 1821 | Sediment | 62°11'51.3420" | 58°59'18.3408" | PQ427691 | PQ427659 | PQ433548 | |||||
| 1870 | Sediment | 62°11'51.3420" | 58°59'18.3408" | PQ427759 | PQ433549 | ||||||||
| Cadophora ramosa | 1845 | Styrofoam | - | - | PQ427705 | PQ427673 | PQ433550 | ||||||
| Cadophora sp.1 | 1810 | Styrofoam | - | - | PQ427756 | ||||||||
| Cadophora sp.2 | 1863 | Soil | 62°13'38.8308" | 58°56'59.0640" | PQ427690 | PQ427679 | |||||||
| Tricladiaceae | Tricladium sp.1 | 1813 | Soil | 62°11'56.83" | 58°59'33.12" | PQ427714 | |||||||
| Tricladium sp.1 | 1855 | Soil | 62°11'56.83" | 58°59'33.12" | PQ427719 | ||||||||
| Tricladium sp.2 | 1840 | - | - | - | PQ427670 | ||||||||
| Tricladium sp.3 | 1854 | Soil | 62°12'12.9708" | 58°57'35.2908" | PQ427776 | PQ427675 | |||||||
| Hypocreales | Cordycipitaceae | Lecanicillium sp. | 1829 | - | - | - | PQ427751 | PQ427663 | |||||
| Hypocreaceae | Hypomyces albidus | 1843 | Fruit body | 62°10'10.8408" | 58°51'02.6460" | PQ427770 | PQ433543 | PQ433539 | |||||
| 1847 | Fruit body | 62°10'10.8408" | 58°51'02.6460" | PQ427766 | PQ433544 | PQ433540 | |||||||
| Nectriaceae | Cosmospora viridescens | 1802 | Moss | 62°13'47.4816" | 58°57'13.3560" | PQ427764 | PQ456771 | ||||||
| Cosmospora sp. | 1834 | Moss | 62°11'47.6700" | 58°58'56.0928" | PQ427768 | PQ456770 | PQ433542 | ||||||
| 1837 | Ice | 62°13'30.5256" | 58°57'31.5036" | PQ427767 | PQ456769 | ||||||||
| 1846 | Moss | 62°13'47.4816" | 58°57'13.3560" | PQ427695 | PQ456766 | ||||||||
| 1852 | Moss | 62°10'10.8408" | 58°51'02.6460" | PQ427726 | PQ456768 | ||||||||
| 1853 | Moss | 62°11'51.3420" | 58°59'18.3408" | PQ427712 | PQ456767 | ||||||||
| Tilachlidiaceae | Psychronectria sp. | 1820 | Moss | 62°09'26.6148" | 58°55'57.0360" | PQ427700 | |||||||
| Ophiocordycipitaceae | Purpureocillium lilacinum | 1902 | - | - | - | PQ427702 | PQ427687 | ||||||
| 1904 | - | - | - | PQ427693 | PQ427688 | ||||||||
| Tolypocladium sp. | 1857 | Macroalga | 62°10'10.8408" | 58°51'02.6460" | PQ427703 | PQ427676 | |||||||
| 1860 | Moss | 62°13'47.4816" | 58°57'13.3560" | PQ427728 | |||||||||
| Mortierellales | Mortierellaceae | Mortierella sp. | 1803 | Fruit body | 62°13'47.4816" | 58°57'13.3560" | PQ427746 | ||||||
| 1875 | Lagoon sediment | 62°12'16.3656" | 58°58'09.4368" | PQ427722 | |||||||||
| 1876 | Styrofoam | - | - | PQ427769 | PQ427684 | ||||||||
| Onygenales | Onygenaceae | Chrysosporium sp. | 1841 | Fruit body | 62°13'47.4816" | 58°57'13.3560" | PQ427698 | PQ427671 | |||||
| Pleosporales | Melanommataceae | Herpotrichia sp.1 | 1808 | Moss | 62°13'47.4816" | 58°57'13.3560" | PQ427761 | PQ427653 | PQ433541 | ||||
| Herpotrichia sp.2 | 1816 | Fruit body | 62°13'47.4816" | 58°57'13.3560" | PQ427774 | PQ427657 | |||||||
| Phaeosphaeriaceae | Phaeosphaeria sp. | 1833 | - | - | - | PQ427665 | |||||||
| Thelebolales | Pseudeurotiaceae | Pseudeurotium sp. | 1850 | Styrofoam | - | - | PQ427745 | PQ427674 | |||||
| 1856 | Moss | 62°12'12.69" | 58°57'36.59" | PQ427706 | |||||||||
| 1861 | Ice | 62°13'30.5256" | 58°57'31.5036" | PQ427771 | |||||||||
| 1865 | Sediment | 62°12'12.69" | 58°57'36.59" | PQ427779 | |||||||||
| 1866 | Moss | 62°15'34.8" | 58°59'11.16" | PQ427778 | PQ427681 | ||||||||
| 1872 | Styrofoam | 62°12'16.3656" | 58°58'09.4368" | PQ427780 | |||||||||
| 1873 | Sediment | 62°09'38.0520" | 58°55'31.8540" | PQ427772 | |||||||||
| 1874 | Sediment | 62°15'34.62" | 58°59'10.56" | PQ427744 | |||||||||
| Pseudogymnoascus appendiculatus | 1828 | Moss | 62°11'51.3420" | 58°59'18.3408" | PQ427731 | PQ427662 | |||||||
| Pseudogymnoascus australis | 1811 | Moss | 62°13'47.4816" | 58°57'13.3560" | PQ427729 | PQ427654 | |||||||
| 1815 | Soil | 62°10'11.4168" | 58°51'10.2096" | PQ427727 | PQ427656 | ||||||||
| Pseudogymnoascus verrucosus | 1801 | Sediment | 62°11'53.5776" | 58°59'38.1156" | PQ427752 | PQ427651 | |||||||
| 1807 | Soil | 62°12'16.3656" | 58°58'09.4368" | PQ427730 | PQ427652 | ||||||||
| 1826 | Soil | 62°12'12.9708" | 58°57'35.2908" | PQ427723 | PQ427661 | ||||||||
| 1832 | River sediment | 62°13'30.2916" | 58°57'14.0508" | PQ427725 | PQ427664 | ||||||||
| 1836 | Soil | 62°09'26.4996" | 58°56'08.3868" | PQ427758 | PQ427667 | ||||||||
| 1864 | Green alga | 62°11'53.8908" | 58°58'16.4640" | PQ427750 | PQ427680 | ||||||||
| Pseudogymnoascus sp.1 | 1814 | Soil | 62°13'48.2520" | 58°57'19.5336" | PQ427711 | PQ427655 | |||||||
| 1817 | Soil | 62°12'12.9708" | 58°57'35.2908" | PQ427720 | PQ427658 | ||||||||
| 1823 | Sediment | 62°11'53.5776" | 58°59'38.1156" | PQ427724 | PQ427660 | ||||||||
| 1835 | River sediment | 62°13'29.1360" | 58°57'09.7236" | PQ427733 | PQ427666 | ||||||||
| 1862 | Sediment | 62°09'38.7468" | 58°55'26.8320" | PQ427749 | PQ427678 | ||||||||
| 1867 | Soil | 62°12'12.9708" | 58°57'35.2908" | PQ427715 | PQ427682 | ||||||||
| 1871 | River sediment | 62°13'30.2916" | 58°57'14.0508" | PQ427718 | PQ427683 | ||||||||
| Pseudogymnoascus sp.2 | 1858 | Soil | 62°12'12.9708" | 58°57'35.2908" | PQ427762 | PQ427677 | |||||||
| Pseudogymnoascus sp.3 | 1838 | Moss | 62°11'51.3420" | 58°59'18.3408" | PQ427775 | PQ427668 |
Table 1.
TOP

