- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- For Contributors
- Policies
- E-Submission

- About
- Browse Articles
-
Special Issues
- Pioneering strategies for overcoming bacterial drug resistance (2026)
- Advancing microbial engineering through synthetic biology (2025)
- Host-associated microbiome (2024)
- Bacterial regulatory mechanisms for the control of complex cellular mechanisms (2023)
- Two years into COVID-19 pandemic: Where are we? (2022)
- Collections
- Policies
- For Contributors
Articles
- Page Path
- HOME > J. Microbiol > Volume 63(3); 2025 > Review
-
Review
Untranslated region engineering strategies for gene overexpression, fine-tuning, and dynamic regulation - Jun Ren†, So Hee Oh†, Dokyun Na*
-
Journal of Microbiology 2025;63(3):e2501033.
DOI: https://doi.org/10.71150/jm.2501033
Published online: March 28, 2025
Department of Biomedical Engineering, Chung-Ang University, Seoul 06974, Republic of Korea
- *Correspondence Dokyun Na blisszen@cau.ac.kr
- †These authors contributed equally to this work.
• Received: January 30, 2025 • Revised: March 8, 2025 • Accepted: March 10, 2025
© The Microbiological Society of Korea
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Supplementary Information
References
Citations
Citations to this article as recorded by 
- Rhodo-Box: A Synthetic Biology Toolbox to Facilitate Metabolic Engineering of Rhodobacter sphaeroides
Matic Kostanjšek, Antoine Raynal, George Dimopoulos, Gerrich Behrendt, Vitor A. P. Martins dos Santos, Jules Beekwilder, Christos Batianis, Ruud A. Weusthuis, Enrique Asin-Garcia, Markus M. M. Bisschops
ACS Synthetic Biology.2026; 15(4): 1400. CrossRef - Production of the recombinant spider silk MaSp2 protein using the marine purple photosynthetic nonsulfur bacterium Rhodovulum sulfidophilum under autotrophic conditions
Miki Suzuki, Keiji Numata
NPG Asia Materials.2026;[Epub] CrossRef - Advancing microbial engineering through synthetic biology
Ki Jun Jeong
Journal of Microbiology.2025; 63(3): e2503100. CrossRef - Recombinase-Mediated Cassette Exchange-Based CRISPR Activation Screening Identifies Hyperosmotic Stress-Resistant Genes in Chinese Hamster Ovary Cells
Minhye Baek, Seokchan Kweon, Yujin Kim, Nathan E. Lewis, Jae Seong Lee, Gyun Min Lee
ACS Synthetic Biology.2025; 14(8): 3116. CrossRef
 ePub Link
ePub Link Cite this Article
Cite this Article
Untranslated region engineering strategies for gene overexpression, fine-tuning, and dynamic regulation

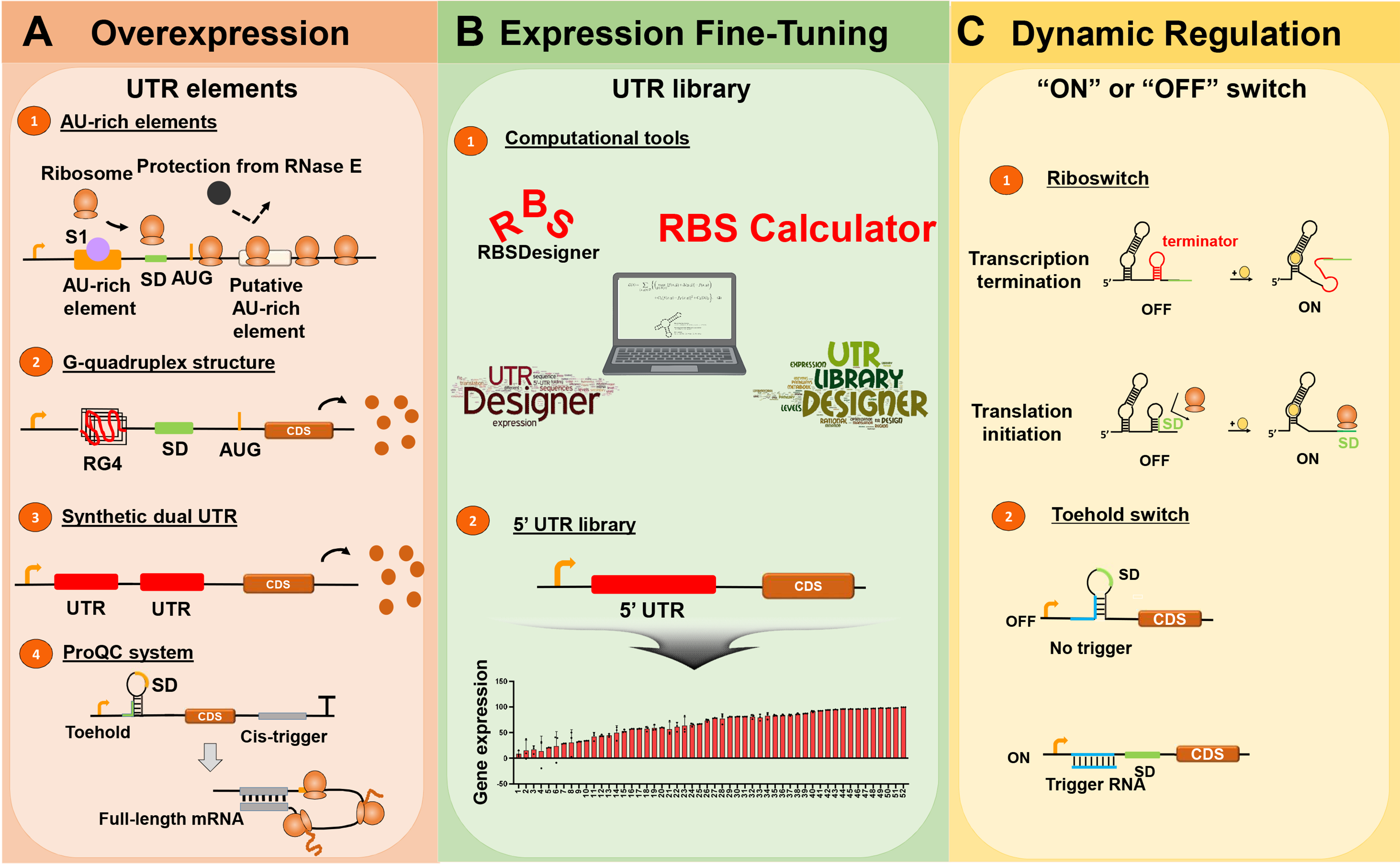
Fig. 1. Schematic illustration on recent UTR engineering strategies.(A) Several strategies utilizing diverse UTR elements, such as AU-rich elements, RG4 structures, synthetic dual UTRs, and the ProQC system, have been employed for overexpression. (B) A 5’UTR library generated using diverse computational tools can enable fine-tuned gene expression. (C) Riboswitches and toehold switches are “on” or “off” switches that dynamically regulate gene expression.
Fig. 1.
Untranslated region engineering strategies for gene overexpression, fine-tuning, and dynamic regulation
| Expression | UTR elements | Description and applications | Ref |
|---|---|---|---|
| Protein over-expression | AU-rich elements | A well-known AU-rich element is originated from sodB gene. S1 protein stabilizes AU-rich mRNAs by recruiting ribosomes and preventing degradation. Knockdown of S1 reduced GFP mRNA by 34% and RFP mRNA by 61%. In its absence, Hfq protein compensates to protect mRNAs from degradation. | |
| RG4 | The enhanced translation by an RG4 structure follows a physical barrier model, where bulky structures in the 5′ UTR guide ribosome movement toward the downstream start codon, increasing translation efficiency. | ||
| Key 5′ UTR features were optimized, including a 25–30 nt spacer between the stabilizer hairpin and RBS and 32% GC content in the spacer. Additional introduction of the RG4 RNA aptamer "Corn" into the 3′ UTR, significantly enhanced protein expression. Employing an optimal 5′ UTR increased recombinant luciferase protein expression by 1.8-fold. Similarly, modifying a gene with both the optimal 5′ UTR motif and the 3′ UTR motif (5×Corn) led to a 3.4-fold increase in recombinant Plasmodium falciparum lactate dehydrogenase (PfLDH) protein expression. | |||
| Synthetic dual UTRs | Two UTRs, enhancing transcription and translation, respectively, were identified from a library of <400,000 randomized UTRs in E. coli and concatenated. This synthetic dual UTR construct enhanced gene expression. The r31n47 dual UTR dramatically enhanced β-lactamase expression, compared with wild-type UTRs. For mCherry, transcript levels, fluorescence intensity, and half-life improved notably. Additionally, this dual UTR enhanced the solubility of both β-lactamase and mCherry proteins, further optimizing protein expression. | ||
| ProQC | The ProQC system was used to express vioB originated from C. violaceum, which converts L-tryptophan into violacein, a polyketide synthase product. This approach enhanced violacein production by 2.3-fold. Additionally, a synthetic fusion enzyme, CrtBI, was expressed using the ProQC system, while geranylgeranyl diphosphate synthase (crtE) was expressed, which resulted in a 2.2-fold increase in lycopene accumulation. | ||
| Expression fine-optimization | RBS | A total of 41 synthetic 5′ UTRs were designed using the RBSCalculator, with translation initiation rates rationally controlled over a 100,000-fold range, enabling precise gene expression regulation. The 5′ UTR_33k_30 exhibited the highest mRNA increasing lacZ by 5-fold, txAbF by 6-fold, and msfGFP by 2.5-fold. | |
| Optimal leader sequence was designed to minimize structure interference between SD and CDS. With the optimal leader sequence, RBS sequences were computationally designed using RBSDesigner to customize gene expression levels. Optimizing the expression levels of key metabolic enzymes encoded by cadA and PP3533 resulted in enhanced metabolite production. The expression-optimized strains produced 2.25 g/L of cadaverine (a 72% increase) and 2.59 g/L of L-proline (a 28% increase). | |||
| The shRBS library achieved a 10⁴-fold dynamic range in expression strength by modifying the spacer regions between the SD sequence and the start codon and in B. licheniformis. To assess the shRBS library for metabolic engineering, five shRBSs were selected to fine-tune leuS gene expression, promoting pulcherriminic acid production in B. licheniformis. Additionally, the rate-limiting enzyme YvmC was overexpressed. The production of pulcherriminic acid varied based on the strength of different shRBSs regulating leuS expression, achieving a two-fold range. The highest production of pulcherriminic acid was observed when U12-9 shRBS was utilized to leuS gene expression. | |||
| A high-throughput screening method using 30–300 bp DNA fragments identified a novel 5′ UTR sequence from the B. subtilis genome, with the highest expression rate. Subsequently, a 5′ UTR library was constructed from the 5′ UTR by introducing mutations to diversify gene expression level. An artificial rib operon in B. subtilisi, in which the genes’ 5′ UTR were replaced with the high-efficient UTR. This artificial rib operon resulted in a 2.09-fold increase in riboflavin production. This enhancement was 4.7-fold higher compared with the operon with its original UTR sequences. | |||
| A library of 90 promoter-UTRs was constructed based on RNA-Seq data analysis. The strongest promoter-UTR sequence, PNCgl1676-UTR, exhibited expression levels over five times higher than Psod-UTR, the most commonly used strong promoter-UTR in C. glutamicum. | |||
| A library of promoter–UTR sequences including RBS from various methanogens was constructed, which achieved a 140-fold dynamic range in expression strength in M. acetivorans. The strongest promoter-RBS was modified to generate six rationally designed high-expression 5′ -UTR variants. When evaluated using β-glucuronidase, they demonstrated a 140-fold range of expression strengths in M. acetivorans. | |||
| 5′ UTR libraries were designed using UTR Library Designer to diversify the expression levels of phlF and mcbR repressors. The expression levels of phlF and mcbR libraries were broadly diversified up to 18.57-fold and 15.14-fold, respectively. The most optimized strain using the libraries achieved a 2.82-fold increase in lycopene production compared with non-optimized strain. Additionally, a strain co-expressing phlF and mcbR with designed UTR sequences exhibited a 16.5-fold increase in 3-HP production compared with the parental strain. | |||
| 5′ UTR libraries of crtE, crtB, and crtI were generated using UTR Library Designer. For each gene, 16 different 5′ UTR sequences were designed. The highest lycopene titer was achieved by expressing crtE, crtB, and crtI, each using its optimally designed UTR sequence. | |||
| To fine-tune pckA expression, four 5′ UTR variants were generated using UTR Library Designer. The best UTR sequence, leading to fine-tuned pckA expression, resulted in a strain that exhibited a 49.8-fold increase in naringenin production compared with the non-optimized strain. | |||
| Dynamic expression regulation | Riboswitch | Riboswitch is a regulatory element in 5’ UTR, controlling the expression of downstream coding sequence due to its structural change upon interaction with its ligand. | |
| Two lysine riboswitches were utilized to activate and repress the expression of aspartate kinase III and homoserine dehydrogenase, respectively, in the lysine-producing strain C. glutamicum OW45. As a result, lysine production increased by 35% in strain QW48 (A263-lysC) and 43% in strain QW54 (R357-hom), compared with the parental strain QW45. | |||
| Toehold switches | Toehold switches are designed to detect specific RNA sequences, known as "triggers," with high sensitivity and specificity. Toehold switch was designed to detect coronavirus RNA, which acts like a trigger. The detection system implemented on a paper-cell free expression system could detect coronavirus. |
| Expression | UTR elements | Advantages | Limitations | Ref |
|---|---|---|---|---|
| Protein Over-expression | AU-rich | Incorporating AU-rich elements into the 5′ UTR region can stabilize mRNA through S1 and Hfq proteins, thereby enhancing protein production. | The relatively long AU-rich element derived from the sodB gene, for example, may enhance RNase E accessibility, potentially reducing mRNA stability. | |
| RG4 | RG4 structures act as internal ribosome entry sites, allowing translation initiation independent of a TSS. Furthermore, their incorporation into the 3′ UTR enhances mRNA stability and protects against RNase-mediated degradation. | The strong helical structure of RG4 complicates the incorporation of other elements like riboswitches into the 5' UTR, because ribonucleotides forming strong intramolecular interactions within the RG4 structure can also interact with other elements, disrupting their structures. | ||
| Synthetic dual UTRs | Dual UTRs elements enhance both transcription and translation with an optimal distance between two UTRs within the 5′ UTR region. | The concatenated UTRs may contain regulatory elements and thus the performance of optimized UTRs may vary under different environmental conditions or growth phases. | ||
| ProQC | The ProQC system ensures translation occurs only in the presence of full-length mRNAs by facilitating circularization through a toehold switch in the 5′ UTR and a cis-trigger sequence in the 3′ UTR. This mechanism enhances full-length translation, thereby improving protein quality in bacteria. | The incorporation of a toehold switch, forming a strong secondary structure, complicates its integration with other 5′ UTR elements. In addition, to further enhance full-length protein production, improving the circularization efficiency and stability of circular mRNA requires further investigation. | ||
| Expression fine-optimization | RBS | A UTR library offers a diverse set of UTR sequences, including RBSs, that can be incorporated into the 5′ UTR region to fine-tune target gene expression by varying ribosome binding affinity. | UTR libraries are designed for static fine-tuning rather than dynamic regulation. | |
| Dynamic expression regulation | Riboswitch | Riboswitches regulate gene expression by binding specific metabolites, switching expression "on" or "off," and controlling downstream gene activity. | Riboswitches may exhibit leaky expression in the OFF state or fail to achieve full activation in the ON state. | |
| Toehold switches | Toehold switches regulate gene expression by binding trigger RNAs, toggling expression "on" or "off," and controlling downstream gene activity. | The strong secondary structure of toehold switch complicates its integration with other 5′ UTR elements, and like riboswitches toehold switches often exhibit leaky expression in the OFF state or fail to achieve full activation in the ON state. Unintended interactions with endogenous RNAs can also lead to undesirable regulatory effects. |
| Software | Description | Ref | Exemplar applications |
|---|---|---|---|
| RBSDesinger | RBSDesigner is a mathematical model accounting for the thermodynamic RBS folding (30 nt from SD) and the interaction of RBS with ribosomes. The thermodynamic parameters in the model were calculated using UNAFold software package. The model’s prediction performance was R2=0.77 – 0.87. | The RBS of the ppc gene, optimized by RBSDesigner, enhanced both rapid cell growth and high 3-AP production in E. coli by facilitating the conversion of phosphoenolpyruvate to oxaloacetate, which is then converted to aspartate and ultimately to 3-AP ( |
|
| URL: |
|||
| RBSCalculator / Operon Calculator | RBSCalculator is a thermodynamic model composed of five energy parameters calculated from -35 to +35 nt of TIR by using NuPACK suite. Its prediction performance was R2=0.51 – 0.95 depending on the origin of UTR. | Optimized expression of fabH and fabZ using RBS sequences designed by RBSCalculator enhanced the synthesis of fatty acyl-ACP from acetyl-CoA. Fatty acyl-ACP was then converted to fatty alcohol by the far gene, resulting in a high production titer of fatty alcohol ( |
|
| Operon Calculator is a biophysical model based on the model of RBSCalculator to predict the translation of bi- and tri-cistronic genes. Its performance is Pearson R2=0.57 – 0.91. | |||
| URL: |
|||
| UTRDesigner / UTR Library Designer | UTRDesigner is a thermodynamic model that accounts for several parameters, calculated from -10 to +35 nt of the TIR using NuPACK. Its performance is R² = 0.81. UTR Library Designer is a tool used to generate UTR sequences with diverse expression levels, utilizing UTRDesigner. | Seo et al. ( |
For enhanced itaconic acid production, multiple genes (acs, gltA, and aceA) involved in acetate assimilation and the glyoxylate shunt pathway were overexpressed using optimized RBS sequences designed by UTRDesigner ( |
| URL: |
Table 1. UTR engineering strategies in bacteria and their applications
Table 2. Advantages and limitations of UTR elements in gene expression modulation
Table 3. Computational tools for predicting and designing UTRs in bacterial gene expression
Table 1.
Table 2.
Table 3.
TOP

